

Sesbania mosaic virus (SeMV) infectious clone: possible mechanism of 3' and 5' end repair and role of polyprotein processing in viral replication
- PMID: 22355344
- PMCID: PMC3280281
- DOI: 10.1371/journal.pone.0031190
Sesbania mosaic virus (SeMV) infectious clone: possible mechanism of 3' and 5' end repair and role of polyprotein processing in viral replication
Abstract
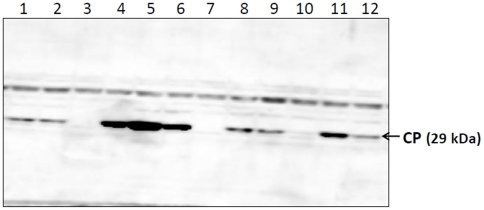
Sesbania mosaic virus (SeMV) is a positive stranded RNA virus belonging to the genus Sobemovirus. Construction of an infectious clone is an essential step for deciphering the virus gene functions in vivo. Using Agrobacterium based transient expression system we show that SeMV icDNA is infectious on Sesbania grandiflora and Cyamopsis tetragonoloba plants. The efficiency of icDNA infection was found to be significantly high on Cyamopsis plants when compared to that on Sesbania grandiflora. The coat protein could be detected within 6 days post infiltration in the infiltrated leaves. Different species of viral RNA (double stranded and single stranded genomic and subgenomic RNA) could be detected upon northern analysis, suggesting that complete replication had taken place. Based on the analysis of the sequences at the genomic termini of progeny RNA from SeMV icDNA infiltrated leaves and those of its 3' and 5' terminal deletion mutants, we propose a possible mechanism for 3' and 5' end repair in vivo. Mutation of the cleavage sites in the polyproteins encoded by ORF 2 resulted in complete loss of infection by the icDNA, suggesting the importance of correct polyprotein processing at all the four cleavage sites for viral replication. Complementation analysis suggested that ORF 2 gene products can act in trans. However, the trans acting ability of ORF 2 gene products was abolished upon deletion of the N-terminal hydrophobic domain of polyprotein 2a and 2ab, suggesting that these products necessarily function at the replication site, where they are anchored to membranes.
Conflict of interest statement
Figures








Similar articles
-
Complete nucleotide sequence of Sesbania mosaic virus: a new virus species of the genus Sobemovirus.Arch Virol. 2001;146(2):209-23. doi: 10.1007/s007050170170. Arch Virol. 2001. PMID: 11315633
-
Functional insights into the role of C-terminal disordered domain of Sesbania mosaic virus RNA-dependent RNA polymerase and the coat protein in viral replication in vivo.Virus Res. 2019 Jul 2;267:26-35. doi: 10.1016/j.virusres.2019.05.003. Epub 2019 May 2. Virus Res. 2019. PMID: 31054934
-
Processing of SeMV polyproteins revisited.Virology. 2010 Jan 5;396(1):106-17. doi: 10.1016/j.virol.2009.09.025. Epub 2009 Oct 25. Virology. 2010. PMID: 19861224
-
Interaction of Sesbania mosaic virus movement protein with the coat protein--implications for viral spread.FEBS J. 2011 Jan;278(2):257-72. doi: 10.1111/j.1742-4658.2010.07943.x. Epub 2010 Dec 1. FEBS J. 2011. PMID: 21122074
-
2A and 2A-like Sequences: Distribution in Different Virus Species and Applications in Biotechnology.Viruses. 2021 Oct 26;13(11):2160. doi: 10.3390/v13112160. Viruses. 2021. PMID: 34834965 Free PMC article. Review.
Cited by
-
Overview on Sobemoviruses and a Proposal for the Creation of the Family Sobemoviridae.Viruses. 2015 Jun 15;7(6):3076-115. doi: 10.3390/v7062761. Viruses. 2015. PMID: 26083319 Free PMC article. Review.
-
Ryegrass mottle virus complete genome determination and development of infectious cDNA by combining two methods- 3' RACE and RNA-Seq.PLoS One. 2023 Dec 5;18(12):e0287278. doi: 10.1371/journal.pone.0287278. eCollection 2023. PLoS One. 2023. PMID: 38051715 Free PMC article.
-
Harnessed viruses in the age of metagenomics and synthetic biology: an update on infectious clone assembly and biotechnologies of plant viruses.Plant Biotechnol J. 2019 Jun;17(6):1010-1026. doi: 10.1111/pbi.13084. Epub 2019 Feb 28. Plant Biotechnol J. 2019. PMID: 30677208 Free PMC article. Review.
-
Plant Viral Proteases: Beyond the Role of Peptide Cutters.Front Plant Sci. 2018 May 17;9:666. doi: 10.3389/fpls.2018.00666. eCollection 2018. Front Plant Sci. 2018. PMID: 29868107 Free PMC article. Review.
-
Groundnut bud necrosis virus encoded NSm associates with membranes via its C-terminal domain.PLoS One. 2014 Jun 11;9(6):e99370. doi: 10.1371/journal.pone.0099370. eCollection 2014. PLoS One. 2014. PMID: 24919116 Free PMC article.
References
-
- Lokesh GL, Gopinath K, Satheshkumar PS, Savithri HS. Complete nucleotide sequence of Sesbania mosaic virus: a new virus species of the genus Sobemovirus. Arch Virol. 2001;146:209–223. - PubMed
-
- Sreenivasalu P, Nayudu MV. Purification and partial characterization of sesbania mosaic virus. Curr Sci. 1982;51:pp.86–87.
-
- Bonneau C, Brugidou C, Chen L, Beachy RN, Fauquet C. Expression of the rice yellow mottle virus P1 protein in vitro and in vivo and its involvement in virus spread. Virology. 1998;244:79–86. - PubMed
-
- Chowdhury SR, Savithri HS. Interaction of Sesbania mosaic virus movement protein with the coat protein–implications for viral spread. FEBS J. 2011;278:257–272. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
