

Replication-competent recombinant porcine reproductive and respiratory syndrome (PRRS) viruses expressing indicator proteins and antiviral cytokines
- PMID: 22355454
- PMCID: PMC3280517
- DOI: 10.3390/v4010102
Replication-competent recombinant porcine reproductive and respiratory syndrome (PRRS) viruses expressing indicator proteins and antiviral cytokines
Abstract
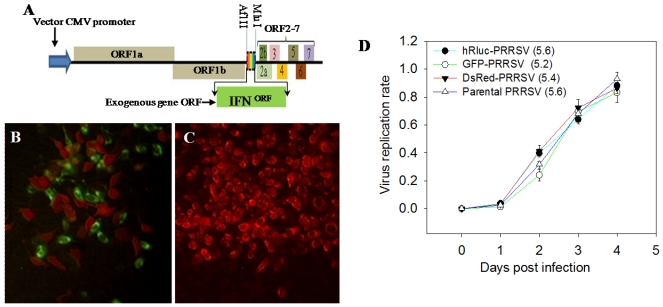
Porcine reproductive and respiratory syndrome virus (PRRSV) can subvert early innate immunity, which leads to ineffective antimicrobial responses. Overcoming immune subversion is critical for developing vaccines and other measures to control this devastating swine virus. The overall goal of this work was to enhance innate and adaptive immunity following vaccination through the expression of interferon (IFN) genes by the PRRSV genome. We have constructed a series of recombinant PRRS viruses using an infectious PRRSV cDNA clone (pCMV-P129). Coding regions of exogenous genes, which included Renilla luciferase (Rluc), green and red fluorescent proteins (GFP and DsRed, respectively) and several interferons (IFNs), were constructed and expressed through a unique subgenomic mRNA placed between ORF1b and ORF2 of the PRRSV infectious clone. The constructs, which expressed Rluc, GFP, DsRed, efficiently produced progeny viruses and mimicked the parental virus in both MARC-145 cells and porcine macrophages. In contrast, replication of IFN-expressing viruses was attenuated, similar to the level of replication observed after the addition of exogenous IFN. Furthermore, the IFN expressing viruses inhibited the replication of a second PRRS virus co-transfected or co-infected. Inhibition by the different IFN subtypes corresponded to their anti-PRRSV activity, i.e., IFNω5 ° IFNα1 > IFN-β > IFNδ3. In summary, the indicator-expressing viruses provided an efficient means for real-time monitoring of viral replication thus allowing high‑throughput elucidation of the role of host factors in PRRSV infection. This was shown when they were used to clearly demonstrate the involvement of tumor susceptibility gene 101 (TSG101) in the early stage of PRRSV infection. In addition, replication‑competent IFN-expressing viruses may be good candidates for development of modified live virus (MLV) vaccines, which are capable of reversing subverted innate immune responses and may induce more effective adaptive immunity against PRRSV infection.
Keywords: host factors; indicator proteins; porcine arterivirus; tumor susceptibility gene 101; type I interferon; virus cDNA infectious clone.
Figures

Similar articles
-
Replication-competent recombinant porcine reproductive and respiratory syndrome (PRRS) virus expressing antiviral cytokine interferon-ω5 as a modified live virus vaccine.Vet Microbiol. 2025 Feb;301:110366. doi: 10.1016/j.vetmic.2025.110366. Epub 2025 Jan 3. Vet Microbiol. 2025. PMID: 39793453
-
Interferon-Induced Transmembrane Protein 3 Is a Virus-Associated Protein Which Suppresses Porcine Reproductive and Respiratory Syndrome Virus Replication by Blocking Viral Membrane Fusion.J Virol. 2020 Nov 23;94(24):e01350-20. doi: 10.1128/JVI.01350-20. Print 2020 Nov 23. J Virol. 2020. PMID: 32999030 Free PMC article.
-
Recombinant Porcine Reproductive and Respiratory Syndrome Virus Expressing Membrane-Bound Interleukin-15 as an Immunomodulatory Adjuvant Enhances NK and γδ T Cell Responses and Confers Heterologous Protection.J Virol. 2018 Jun 13;92(13):e00007-18. doi: 10.1128/JVI.00007-18. Print 2018 Jul 1. J Virol. 2018. PMID: 29643245 Free PMC article.
-
Innate and adaptive immunity against Porcine Reproductive and Respiratory Syndrome Virus.Vet Immunol Immunopathol. 2015 Sep 15;167(1-2):1-14. doi: 10.1016/j.vetimm.2015.07.003. Epub 2015 Jul 17. Vet Immunol Immunopathol. 2015. PMID: 26209116 Free PMC article. Review.
-
Live porcine reproductive and respiratory syndrome virus vaccines: Current status and future direction.Vaccine. 2015 Aug 7;33(33):4069-80. doi: 10.1016/j.vaccine.2015.06.092. Epub 2015 Jul 4. Vaccine. 2015. PMID: 26148878 Review.
Cited by
-
A Recombinant Porcine Reproductive and Respiratory Syndrome Virus Stably Expressing DsRed Protein Based on Bacterial Artificial Chromosome System.Front Microbiol. 2022 Jan 21;13:839845. doi: 10.3389/fmicb.2022.839845. eCollection 2022. Front Microbiol. 2022. PMID: 35126342 Free PMC article.
-
Antiviral regulation in porcine monocytic cells at different activation states.J Virol. 2014 Oct;88(19):11395-410. doi: 10.1128/JVI.01714-14. Epub 2014 Jul 23. J Virol. 2014. PMID: 25056886 Free PMC article.
-
HRS Facilitates Newcastle Disease Virus Replication in Tumor Cells by Promoting Viral Budding.Int J Mol Sci. 2024 Sep 19;25(18):10060. doi: 10.3390/ijms251810060. Int J Mol Sci. 2024. PMID: 39337546 Free PMC article.
-
Tumor Susceptibility Gene 101 (TSG101) Contributes to Virion Formation of Porcine Reproductive and Respiratory Syndrome Virus via Interaction with the Nucleocapsid (N) Protein along with the Early Secretory Pathway.J Virol. 2022 Mar 23;96(6):e0000522. doi: 10.1128/jvi.00005-22. Epub 2022 Jan 26. J Virol. 2022. PMID: 35080428 Free PMC article.
-
Genome-wide analysis of antiviral signature genes in porcine macrophages at different activation statuses.PLoS One. 2014 Feb 5;9(2):e87613. doi: 10.1371/journal.pone.0087613. eCollection 2014. PLoS One. 2014. PMID: 24505295 Free PMC article.
References
-
- Benfield D.A., Nelson E., Collins J.E., Harris L., Goyal S.M., Robison D., Christianson W.T., Morrison R.B., Gorcyca D., Chladek D. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet. Diagn. Invest. 1992;4:127–133. - PubMed
-
- Collins J.E., Benfield D.A., Christianson W.T., Harris L., Hennings J.C., Shaw D.P., Goyal S.M., McCullough S., Morrison R.B., Joo H.S., et al. Isolation of swine infertility and respiratory syndrome virus (isolate ATCC VR-2332) in North America and experimental reproduction of the disease in gnotobiotic pigs. J. Vet. Diagn. Invest. 1992;4:117–126. doi: 10.1177/104063879200400201. - DOI - PubMed
-
- Neumann E.J., Kliebenstein J.B., Johnson C.D., Mabry J.W., Bush E.J., Seitzinger A.H., Green A.L., Zimmerman J.J. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J. Am. Vet. Med. Assoc. 2005;227:385–392. - PubMed
-
- Meulenberg J.J. PRRSV, the virus. Vet. Res. 2000;31:11–21. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
