

Multibudded tubules formed by COPII on artificial liposomes
- PMID: 22355536
- PMCID: PMC3216505
- DOI: 10.1038/srep00017
Multibudded tubules formed by COPII on artificial liposomes
Abstract
COPII-coated vesicles form at the endoplasmic reticulum for cargo transport to the Golgi apparatus. We used in vitro reconstitution to examine the roles of the COPII scaffold in remodeling the shape of a lipid bilayer. Giant Unilamellar Vesicles were examined using fast confocal fluorescence and cryo-electron microscopy in order to avoid separation steps and minimize mechanical manipulation. COPII showed a preference for high curvature structures, but also sufficient flexibility for binding to low curvatures. The COPII proteins induced beads-on-a-string-like constricted tubules, similar to those previously observed in cells. We speculate about a mechanical pathway for vesicle fission from these multibudded COPII-coated tubules, considering the possibility that withdrawal of the Sar1 amphipathic helix upon GTP hydrolysis leads to lipid bilayer destabilization resulting in fission.
Figures

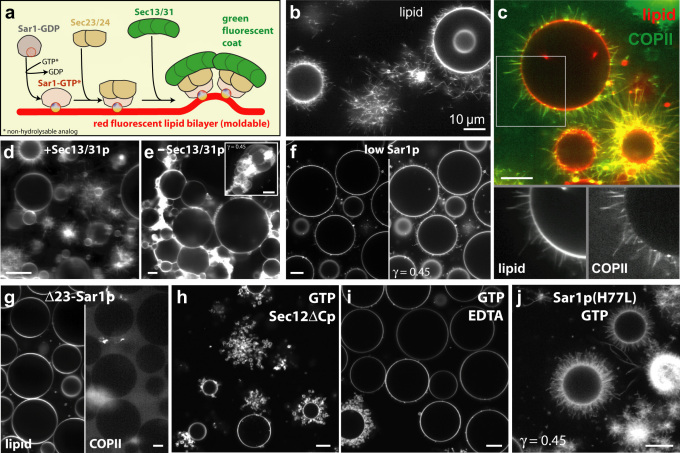
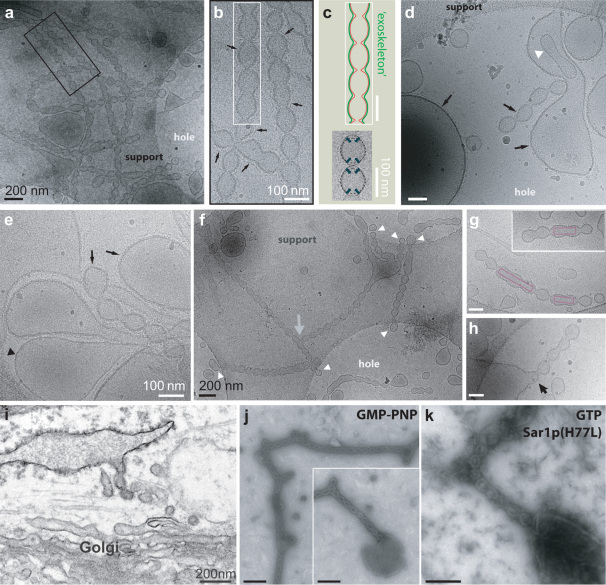
References
-
- Lee M. C. et al. Sar1p N-terminal helix initiates membrane curvature and completes the fission of a COPII vesicle. Cell 122, 605–617 (2005). - PubMed
-
- Saito Y., Kimura K., Oka T. & Nakano A. Activities of mutant Sar1 proteins in guanine nucleotide binding, GTP hydrolysis, and cell-free transport from the endoplasmic reticulum to the Golgi apparatus. J. Biochem. 124, 816–823 (1998). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
