

Potential role of LMP2 as tumor-suppressor defines new targets for uterine leiomyosarcoma therapy
- PMID: 22355695
- PMCID: PMC3240965
- DOI: 10.1038/srep00180
Potential role of LMP2 as tumor-suppressor defines new targets for uterine leiomyosarcoma therapy
Abstract
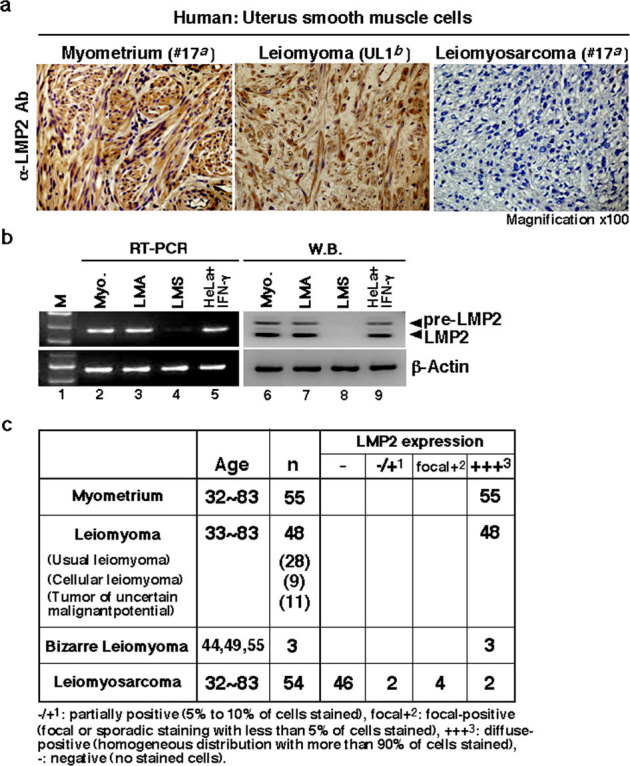
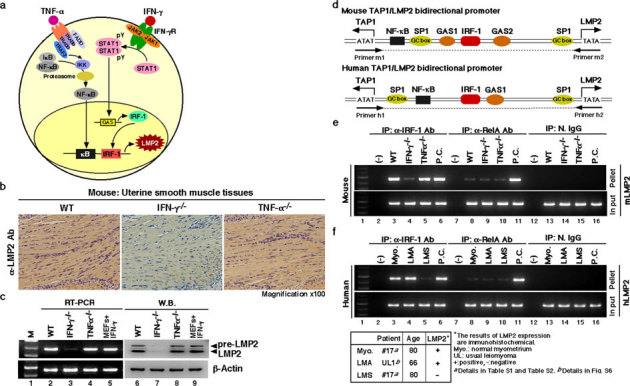
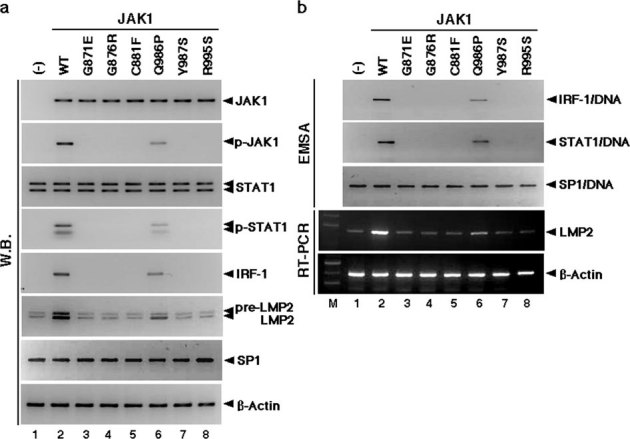
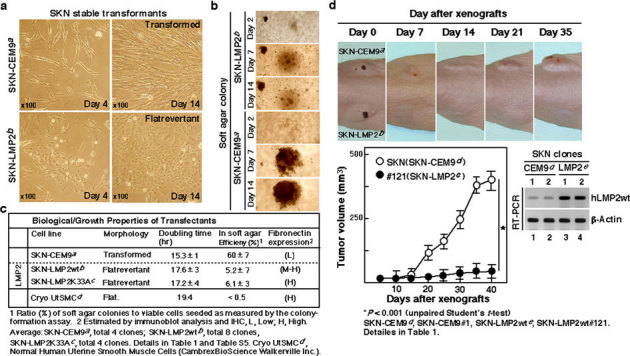
Although the majority of smooth muscle neoplasms found in the uterus are benign, uterine leiomyosarcoma (LMS) is extremely malignant, with high rates of recurrence and metastasis. We earlier reported that mice with a homozygous deficiency for LMP2, an interferon (IFN)-γ-inducible factor, spontaneously develop uterine LMS. The IFN-γ pathway is important for control of tumor growth and invasion and has been implicated in several cancers. In this study, experiments with human and mouse uterine tissues revealed a defective LMP2 expression in human uterine LMS that was traced to the IFN-γ pathway and the specific effect of JAK-1 somatic mutations on the LMP2 transcriptional activation. Furthermore, analysis of a human uterine LMS cell line clarified the biological significance of LMP2 in malignant myometrium transformation and cell cycle, thus implicating LMP2 as an anti-tumorigenic candidate. This role of LMP2 as a tumor suppressor may lead to new therapeutic targets in human uterine LMS.
Figures
References
-
- Zaloudek C. & Hendrickson M. R. Mesenchymal tumors of the uterus, in Kurman RJ. (ed): Blaustein's Pathology of the Female Genital Tract (ed 5). New York, Springer-Verlag 561–578 (2002).
-
- Gadducci A., et al. Uterine leiomyosarcoma: analysis of treatment failures and survival. Gynecol. Oncol. 62, 25–32 (1996). - PubMed
-
- Nordal R. & Thoresen S. Uterine sarcomas in Norway 1956–1992: incidence, survival and mortality. Eur. J. Cancer 33, 907–311 (1997). - PubMed
-
- Brooks S. E., Zhan M., Cote T. & Baquet C. R. Surveillance, epidemiology, and end results analysis of 2677 cases of uterine sarcoma 1989–1999. Gynecol. Oncol. 93, 204–208 (2004). - PubMed
-
- Dusenbery K. E., Potish R. A., Argenta P. A. & Judson P. L. On the apparent failure of adjuvant pelvic radiotherapy to improve survival for women with uterine sarcomas confined to the uterus. Am. J. Clin. Oncol. 28, 295–300 (2005). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
