

NLRP5 mediates mitochondrial function in mouse oocytes and embryos
- PMID: 22357545
- PMCID: PMC3364921
- DOI: 10.1095/biolreprod.111.093583
NLRP5 mediates mitochondrial function in mouse oocytes and embryos
Abstract
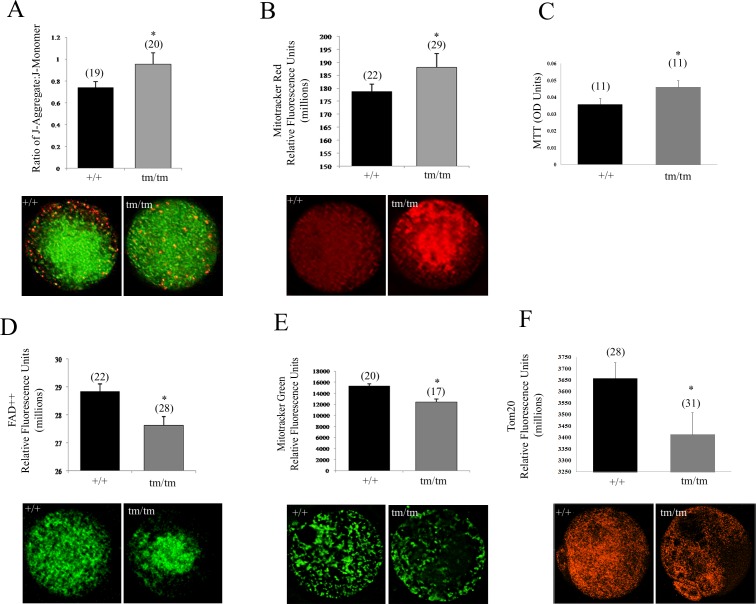
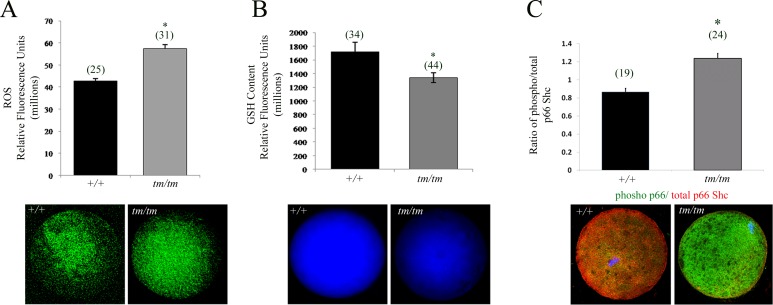
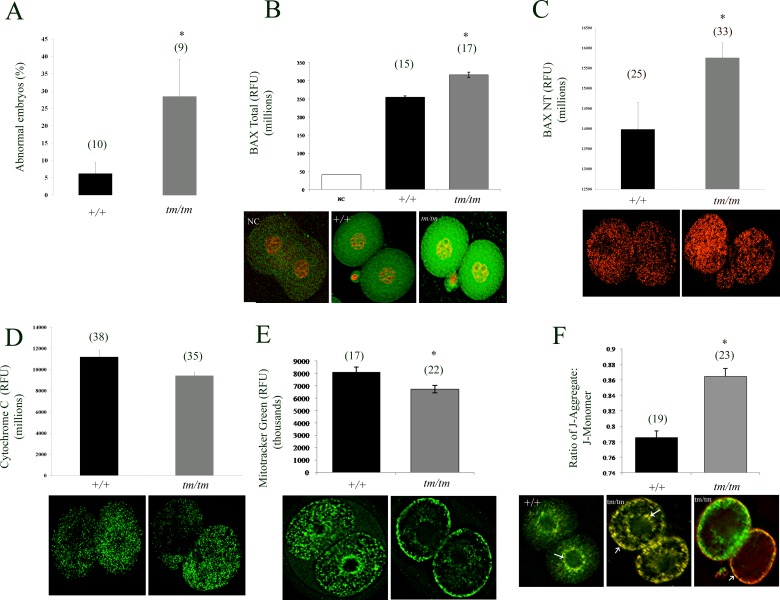
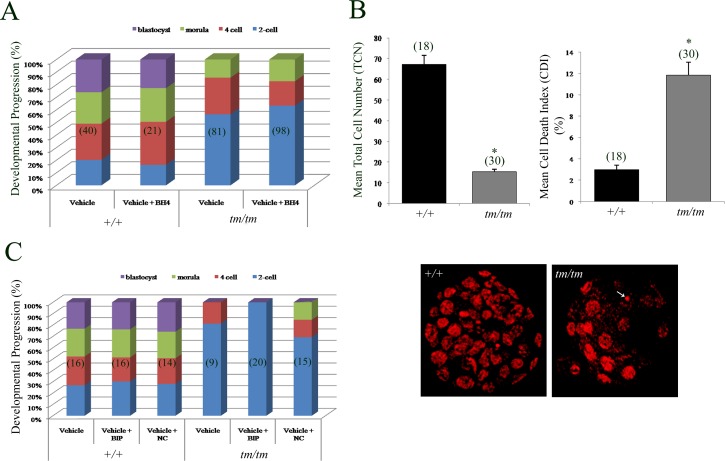
Unraveling molecular pathways responsible for regulation of early embryonic development is crucial for our understanding of female infertility. Maternal determinants that control the transition from oocyte to embryo are crucial molecules that govern developmental competence of the newly conceived zygote. We describe a series of defects that are triggered by a disruption of maternal lethal effect gene, Nlrp5. Previous studies have shown that Nlrp5 hypomorph embryos fail to develop beyond the two-cell stage. Despite its importance in preimplantation development, the mechanism by which the embryo arrest occurs remains unclear. We confirmed that Nlrp5 mutant and wild-type females possess comparable ovarian germ pool and follicular recruitment rates. However, ovulated oocytes lacking Nlrp5 have abnormal mitochondrial localization and increased activity in order to sustain physiological ATP content. This results in an accumulation of reactive oxygen species and increased cellular stress causing mitochondrial depletion. Compromised cellular state is also accompanied by increased expression of cell death inducer Bax and depletion of cytochrome c. However, neither genetic deletion (Bax/Nlrp5 double knockout) nor mimetic interference (BH4 domain or Bax inhibitory peptide) were sufficient to alleviate embryo demise caused by depletion of Nlrp5. We therefore conclude that lack of Nlrp5 in oocytes triggers premature activation of the mitochondrial pool, causing mitochondrial damage that cannot be rescued by inactivation of Bax.
Figures
Similar articles
-
Maternal depletion of NLRP5 blocks early embryogenesis in rhesus macaque monkeys (Macaca mulatta).Hum Reprod. 2009 Feb;24(2):415-24. doi: 10.1093/humrep/den403. Epub 2008 Dec 3. Hum Reprod. 2009. PMID: 19054779
-
Knockdown of NLRP5 arrests early embryogenesis in sows.Anim Reprod Sci. 2015 Dec;163:151-6. doi: 10.1016/j.anireprosci.2015.11.004. Epub 2015 Nov 10. Anim Reprod Sci. 2015. PMID: 26585895
-
Maternally derived FILIA-MATER complex localizes asymmetrically in cleavage-stage mouse embryos.Development. 2008 Jan;135(2):259-69. doi: 10.1242/dev.011445. Epub 2007 Dec 5. Development. 2008. PMID: 18057100
-
Mitochondrial function in the human oocyte and embryo and their role in developmental competence.Mitochondrion. 2011 Sep;11(5):797-813. doi: 10.1016/j.mito.2010.09.012. Epub 2010 Oct 7. Mitochondrion. 2011. PMID: 20933103 Review.
-
Mitochondrial unfolded protein response: a stress response with implications for fertility and reproductive aging.Fertil Steril. 2019 Feb;111(2):197-204. doi: 10.1016/j.fertnstert.2018.11.048. Fertil Steril. 2019. PMID: 30691623 Review.
Cited by
-
Fam70A binds Wnt5a to regulate meiosis and quality of mouse oocytes.Cell Prolif. 2020 Jun;53(6):e12825. doi: 10.1111/cpr.12825. Epub 2020 May 11. Cell Prolif. 2020. PMID: 32391621 Free PMC article.
-
Unsolved Mysteries in NLR Biology.Front Immunol. 2013 Sep 17;4:285. doi: 10.3389/fimmu.2013.00285. Front Immunol. 2013. PMID: 24062750 Free PMC article. Review.
-
Single-nucleotide polymorphisms in FLT3, NLRP5, and TGIF1 are associated with litter size in Small-tailed Han sheep.Arch Anim Breed. 2021 Dec 17;64(2):475-486. doi: 10.5194/aab-64-475-2021. eCollection 2021. Arch Anim Breed. 2021. PMID: 35024433 Free PMC article.
-
Totipotency: what it is and what it is not.Stem Cells Dev. 2014 Apr 15;23(8):796-812. doi: 10.1089/scd.2013.0364. Epub 2014 Feb 12. Stem Cells Dev. 2014. PMID: 24368070 Free PMC article.
-
NLRP7 and the Genetics of Hydatidiform Moles: Recent Advances and New Challenges.Front Immunol. 2013 Aug 20;4:242. doi: 10.3389/fimmu.2013.00242. eCollection 2013. Front Immunol. 2013. PMID: 23970884 Free PMC article.
References
-
- Tong ZB, Gold L, Pfeifer KE, Dorward H, Lee E, Bondy CA, Dean J, Nelson LM. Mater, a maternal effect gene required for early embryonic development in mice. Nat Genet 2000; 26: 267 268 - PubMed
-
- Christians E, Davis AA, Thomas SD, Benjamin IJ. Maternal effect of Hsf1 on reproductive success. Nature 2000; 407: 693 694 - PubMed
-
- Tsukamoto S, Kuma A, Mizushima N. The role of autophagy during the oocyte-to-embryo transition. Autophagy 2008; 4: 1076 1078 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
