

Passive resting state and history of antagonist muscle activity shape active extensions in an insect limb
- PMID: 22357791
- PMCID: PMC3362280
- DOI: 10.1152/jn.01072.2011
Passive resting state and history of antagonist muscle activity shape active extensions in an insect limb
Abstract
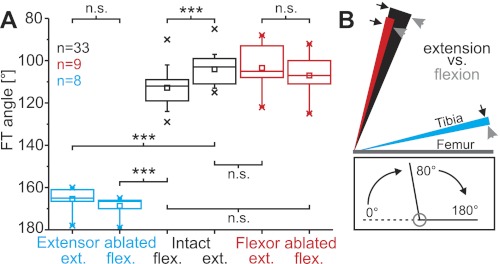
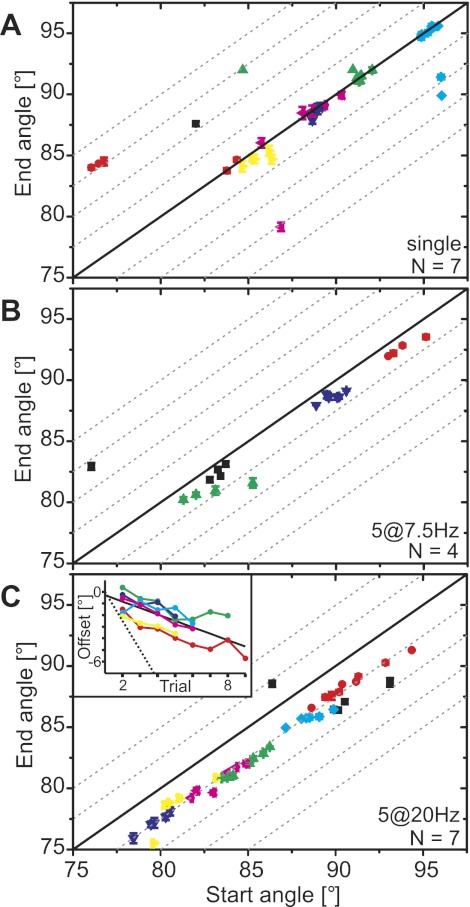
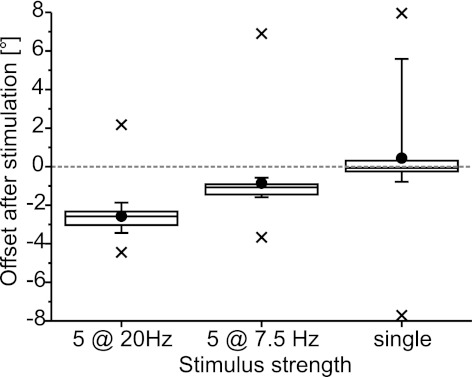
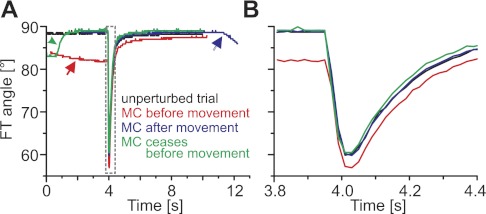
Limb movements can be driven by muscle contractions, external forces, or intrinsic passive forces. For lightweight limbs like those of insects or small vertebrates, passive forces can be large enough to overcome the effects of gravity and may even generate limb movements in the absence of active muscle contractions. Understanding the sources and actions of such forces is therefore important in understanding motor control. We describe passive properties of the femur-tibia joint of the locust hind leg. The resting angle is determined primarily by passive properties of the relatively large extensor tibiae muscle and is influenced by the history of activation of the fast extensor tibiae motor neuron. The resting angle is therefore better described as a history-dependent resting state. We selectively stimulated different flexor tibiae motor neurons to generate a range of isometric contractions of the flexor tibiae muscle and then stimulated the fast extensor tibiae motor neuron to elicit active tibial extensions. Residual forces in the flexor muscle have only a small effect on subsequent active extensions, but the effect is larger for distal than for proximal flexor motor neurons and varies with the strength of flexor activation. We conclude that passive properties of a lightweight limb make substantial and complex contributions to the resting state of the limb that must be taken into account in the patterning of neuronal control signals driving its active movements. Low variability in the effects of the passive forces may permit the nervous system to accurately predict their contributions to behavior.
Figures
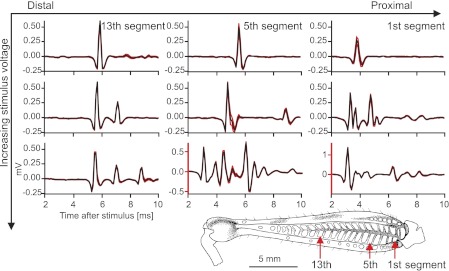
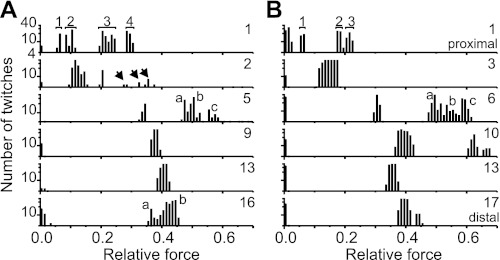
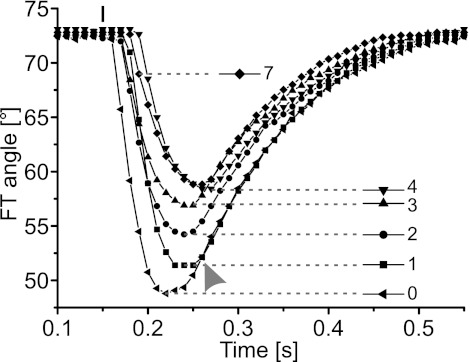
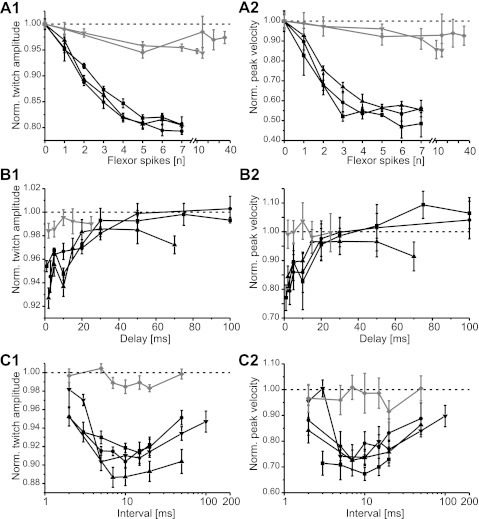
References
-
- Ache JM. From Spike to Movement—Biomechanics and Passive Forces in an Insect Joint (MSc thesis). Cologne, Germany: University of Cologne, 2010
-
- Ahn AN, Full RJ. A motor and a brake: two leg extensor muscles acting at the same joint manage energy differently in a running insect. J Exp Biol 205: 379–389, 2002 - PubMed
-
- Ball EE, Goodman CS. Muscle development in the grasshopper embryo. II. Syncytial origin of the extensor tibiae muscle pioneers. Dev Biol 111: 399–416, 1985a - PubMed
-
- Ball EE, Goodman CS. Muscle development in the grasshopper embryo. III. Sequential origin of the flexor tibiae muscle pioneers. Dev Biol 111: 417–424, 1985b - PubMed
-
- Bässler D, Büschges A, Meditz S, Bässler U. Correlation between muscle structure and filter characteristics of the muscle-joint system in three orthopteran insect species. J Exp Biol 199: 2169–2183, 1996 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources