

Emerging regulatory mechanisms in ubiquitin-dependent cell cycle control
- PMID: 22357967
- PMCID: PMC3283867
- DOI: 10.1242/jcs.091199
Emerging regulatory mechanisms in ubiquitin-dependent cell cycle control
Abstract
The covalent modification of proteins with ubiquitin is required for accurate cell division in all eukaryotes. Ubiquitylation depends on an enzymatic cascade, in which E3 enzymes recruit specific substrates for modification. Among ~600 human E3s, the SCF (Skp1-cullin1-F-box) and the APC/C (anaphase-promoting complex/cyclosome) are known for driving the degradation of cell cycle regulators to accomplish irreversible cell cycle transitions. The cell cycle machinery reciprocally regulates the SCF and APC/C through various mechanisms, including the modification of these E3s or the binding of specific inhibitors. Recent studies have provided new insight into the intricate relationship between ubiquitylation and the cell division apparatus as they revealed roles for atypical ubiquitin chains, new mechanisms of substrate and E3 regulation, as well as extensive crosstalk between ubiquitylation enzymes. Here, we review these emerging regulatory mechanisms of ubiquitin-dependent cell cycle control and discuss how their manipulation might provide therapeutic benefits in the future.
Figures
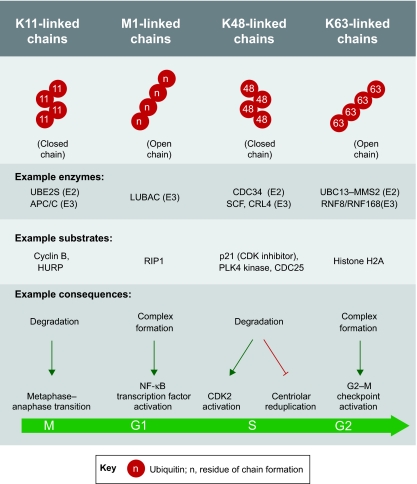
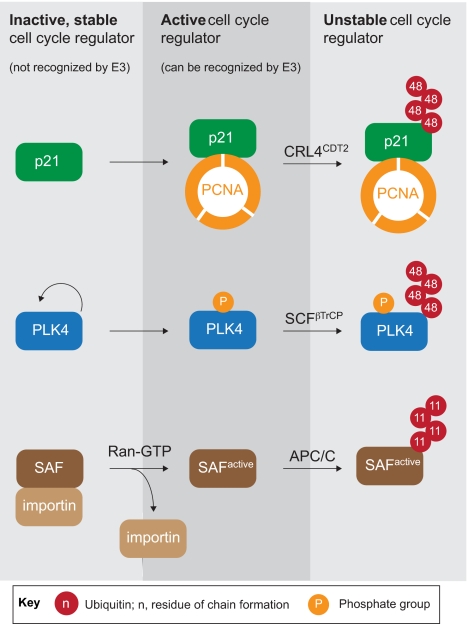
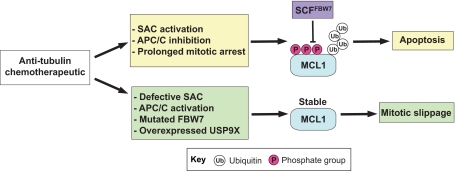
Comment on
- J Cell Sci. 125(2):249.
- J Cell Sci. 125(2):265.
- J Cell Sci. 125(2):277.
References
-
- Arias E. E., Walter J. C. (2006). PCNA functions as a molecular platform to trigger Cdt1 destruction and prevent re-replication. Nat. Cell Biol. 8, 84-90 - PubMed
-
- Baboshina O. V., Haas A. L. (1996). Novel multiubiquitin chain linkages catalyzed by the conjugating enzymes E2EPF and RAD6 are recognized by 26 S proteasome subunit 5. J. Biol. Chem. 271, 2823-2831 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
