

Review
doi: 10.1038/nrm3289.
Y-family DNA polymerases and their role in tolerance of cellular DNA damage
Affiliations
- PMID: 22358330
- PMCID: PMC3630503
- DOI: 10.1038/nrm3289
Item in Clipboard
Review
Y-family DNA polymerases and their role in tolerance of cellular DNA damage
Nat Rev Mol Cell Biol.
.
Abstract
The past 15 years have seen an explosion in our understanding of how cells replicate damaged DNA and how this can lead to mutagenesis. The Y-family DNA polymerases lie at the heart of this process, which is commonly known as translesion synthesis. This family of polymerases has unique features that enable them to synthesize DNA past damaged bases. However, as they exhibit low fidelity when copying undamaged DNA, it is essential that they are only called into play when they are absolutely required. Several layers of regulation ensure that this is achieved.
Figures
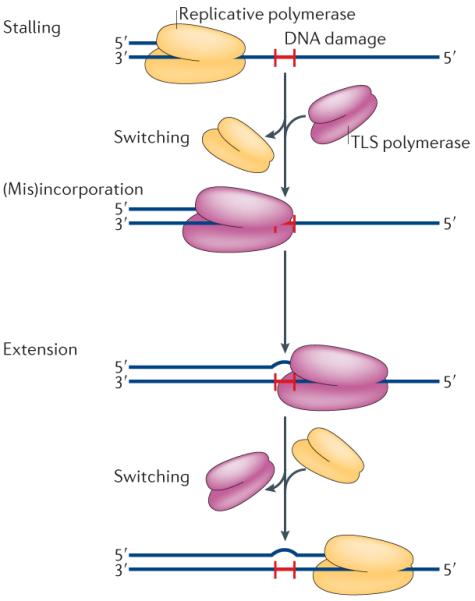
Translesion synthesis (TLS) is a multi-step process. The replicative DNA polymerase stalls at a site of DNA damage (red `H'), then a TLS polymerase (or polymerases) is recruited to the primer terminus and correct, or incorrect, bases are inserted opposite the lesion. The inserted base is subsequently extended to complete TLS. All TLS polymerases are distributive and dissociate from the primer terminus after synthesizing a short tract of DNA. They are then quickly replaced by the cell's replicative polymerase, which completes genome duplication.
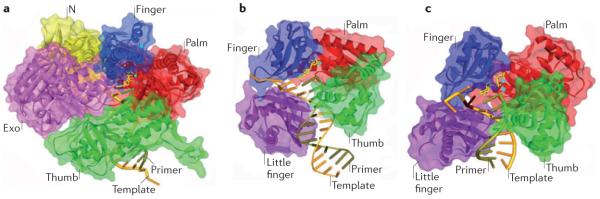
a | Structure of the B-family DNA polymerase from the bacteriophage RB69, with DNA and the incoming dCTP opposite dG (Protein Data Bank (PDB) code 3NCI) shown for comparison to the Y-family structures. The main domains are colour coded (palm = red, thumb = green, finger = blue, exonuclease (Exo) = purple, amino-terminal domain (N) = yellow). b | Crystal structure of the Sulfolobus solfataricus Y-family polymerase Dpo4 in a ternary complex with DNA and incoming ddADP (PDB code 1JX4). The additional little-finger domain is shown in purple. Note the absence of an exonuclease domain. c | Crystal structure of the human Y-family DNA polymerase Pol η in a ternary complex with a cyclobutane pyrimidine dimer (CPD). In this view, the 3′T of the CPD is in the active site and is correctly paired with incoming dATP (PDB code 3MR3). The template strand is shown in rust colour, and the primer is olive green. The incoming dNTP is shown in yellow. The burgundy stick represents the position of the CPD. The small blue spheres represent the metal ions. The protein backbone is represented by ribbon surrounded by semi-transparent solvent accessible surface. This figure was kindly created by A. Vaisman (US National Institute of Child Health and Human Development/US National Institutes of Health) using the Discovery Studio Visualizer program.
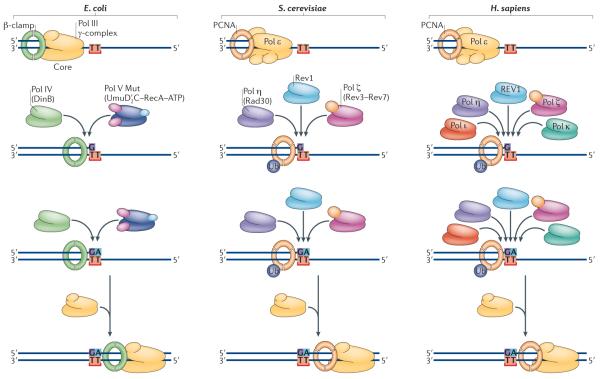
Replicative polymerases, such as Escherichia coli DNA polymerase III (Pol III; which is made up of a core, β-clamp and γ-complex) or eukaryotic Pol ε, stall at the site of a DNA lesion (represented in the figure as TT). Translesion synthesis (TLS) polymerases are recruited to the site via interactions with the replicative sliding processivity clamp (the β-clamp in E. coli and PCNA in eukaryotes). E. coli has a choice of two Y-family polymerases, Pol IV (also known as DinB) and Pol V Mut (which is comprised of UmuD′2 C–RecA–ATP). Saccharomyces cerevisiae also has a choice of two Y-family polymerases, Rev1 and Pol η (which is the product of the RAD30 gene), as well as the B-family TLS polymerase Pol ζ (which is a heterodimer of the Rev3 and Rev7 proteins). In humans, at least five TLS polymerases can be recruited to sites of arrested replication, including Pol η, Pol ι, Pol κ, REV1 and Pol ζ. The likelihood that this insertion step is error-prone, as shown for E. coli and S. cerevisiae, or error-free, as shown for humans, will depend on the nature of the DNA lesion and the polymerase utilized. The extension step may be facilitated by the same enzyme that performed the (mis)insertion or by a completely different polymerase. Once the nascent DNA chain has been extended beyond the lesion, the TLS polymerase is replaced by the cell's replicative DNA polymerase.
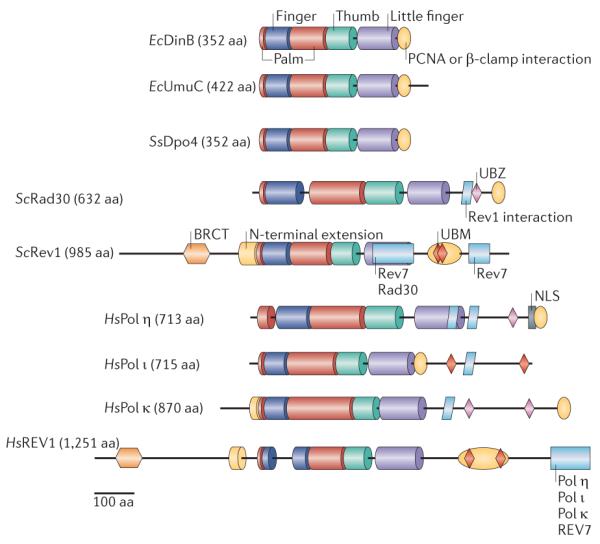
The active sites of the polymerases (Pols) are contained within the palm, thumb, finger and little-finger domains, colour coded as in FIG. 2. The additional domains involved in localization and regulation are BRCT (breast cancer-associated protein-1 carboxy-terminal domain), NLS (nuclear localization signal) and UBM and UBZ (ubiquitin-binding motifs). Species abbreviations: Ec, Escherichia coli; Hs, Homo sapiens; Sc, Saccharomyces cerevisiae; Ss, Sulfolobus solfataricus. aa, amino acid.
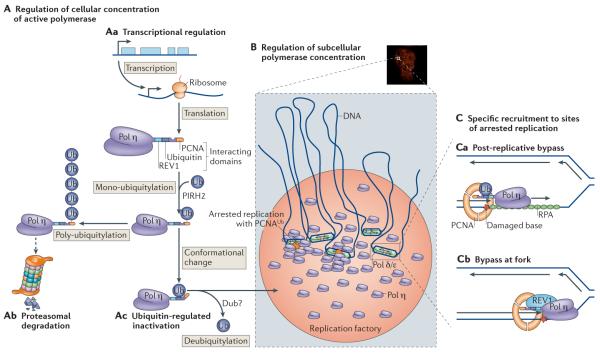
A | Different ways of regulating the cellular concentrations of DNA polymerase η (Pol η). This can be achieved by regulation of transcription (Aa) and subsequent mono-ubiquitylation by PIRH2 (REF. 86), which can lead to poly-ubiquitylation and degradation by the proteasome (Ab) or to inactivation following a conformational change, (Ac). The ubiquitin (Ub) can be removed by a de-ubiquitinase enzyme (Dub). B | Accumulation in replication factories. A nucleus in which Pol η is localized in replication factories is shown (top) with a schematic of polymerases and replication forks that are present in the factories (bottom). C | Recruitment of Pol η by ubiquitylated PCNA at a gap behind the fork (Ca) or by REV1 close to the fork (Cb).
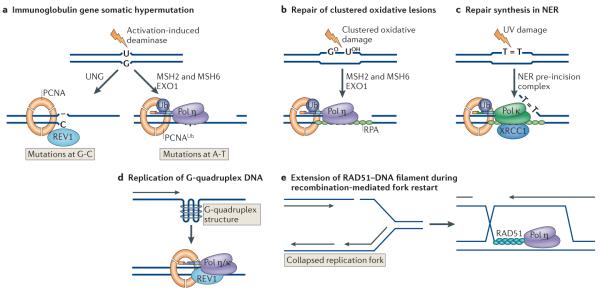
a | The first step in somatic hypermutation at immunoglobulin loci is the deamination of cytosine to uracil by activation-induced deaminase. Removal of uracil by the uracil DNA glycosylase (UNG) generates an abasic site. Replication past the abasic site can result in REV1 inserting C, generating a C-G to G-C transversion. An alternative process involving the mismatch repair proteins MSH2 and MSH6 and exonuclease 1 (EXO1) leads to the formation of a single-strand gap, ubiquitylation of PCNA and recruitment of DNA polymerase η (Pol η), generating mutations at A-T base pairs. b | In a possibly related process, Pol η, recruited by ubiquitylated PCNA, is required for synthesis of gaps generated on removal of clustered oxidative damage (GO, 8-oxoguanine; UOH, 5-hydroxyuracil), in a process that also involves MSH2, MSH6 and EXO1. c | Repair synthesis during nucleotide excision repair (NER) is partially dependent on Pol κ being recruited to the repair site by ubiquitylated PCNA and the scaffold protein XRCC1 (XPG is a nuclease that cleaves 3′ to the damage). d | Y-family polymerases are needed to replicate unusual structures such as G quadruplexes in DNA. e | Homologous recombination involves strand invasion mediated by RAD51 and the formation of a D-loop. Extension of the invading strand in an in vitro system requires Pol η.
References
-
- Ohmori H, et al. The Y-family of DNA polymerases. Mol. Cell. 2001;8:7–8. - PubMed
-
- Rupp WD, Howard-Flanders P. Discontinuities in the DNA synthesized in an excision-defective strain of Escherichia coli following ultraviolet irradiation. J. Mol. Biol. 1968;31:291–304. - PubMed
-
First demonstration of gaps in newly synthesized DNA in E. coli.
-
- Radman M. In: Molecular and Environmental Aspects of Mutagenesis. Prakash L, Sherman F, Miller MW, Lawrence CW, Tabor HW, editors. Charles, C. Thomas; Springfield, Illinois: 1974. pp. 128–142.
-
- Kato T, Shinoura Y. Isolation and characterization of mutants of Escherichia coli deficient in induction of mutations by ultraviolet light. Mol. Gen. Genet. 1977;156:121–131. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
