

Borrelia burgdorferi requires the alternative sigma factor RpoS for dissemination within the vector during tick-to-mammal transmission
- PMID: 22359504
- PMCID: PMC3280991
- DOI: 10.1371/journal.ppat.1002532
Borrelia burgdorferi requires the alternative sigma factor RpoS for dissemination within the vector during tick-to-mammal transmission
Abstract
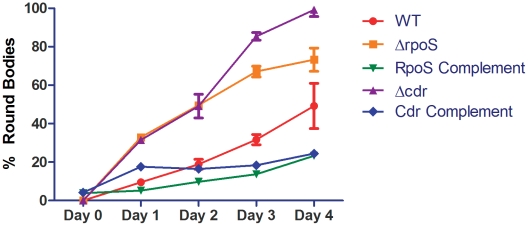
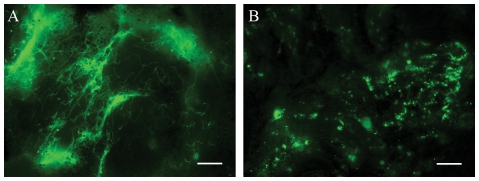
While the roles of rpoS(Bb) and RpoS-dependent genes have been studied extensively within the mammal, the contribution of the RpoS regulon to the tick-phase of the Borrelia burgdorferi enzootic cycle has not been examined. Herein, we demonstrate that RpoS-dependent gene expression is prerequisite for the transmission of spirochetes by feeding nymphs. RpoS-deficient organisms are confined to the midgut lumen where they transform into an unusual morphotype (round bodies) during the later stages of the blood meal. We show that round body formation is rapidly reversible, and in vitro appears to be attributable, in part, to reduced levels of Coenzyme A disulfide reductase, which among other functions, provides NAD+ for glycolysis. Our data suggest that spirochetes default to an RpoS-independent program for round body formation upon sensing that the energetics for transmission are unfavorable.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

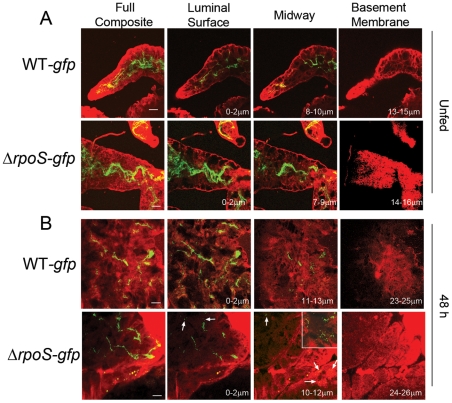
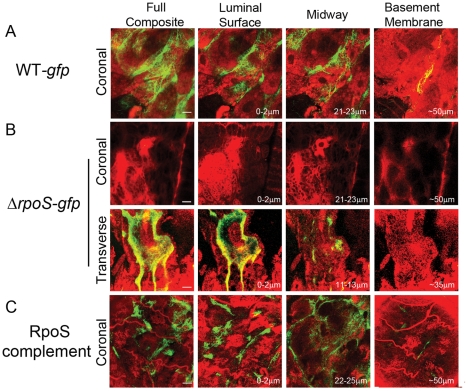
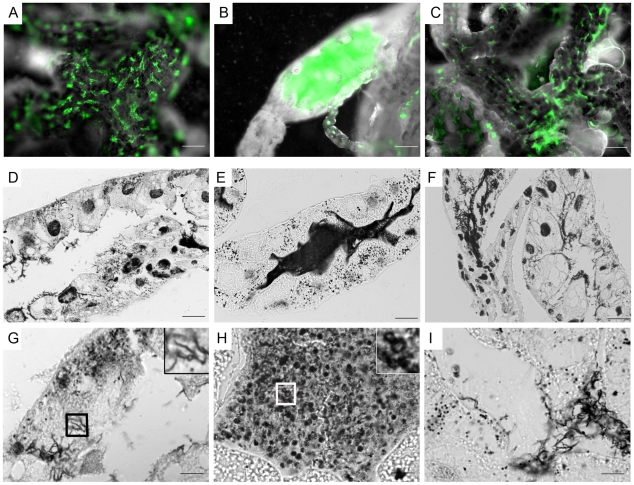
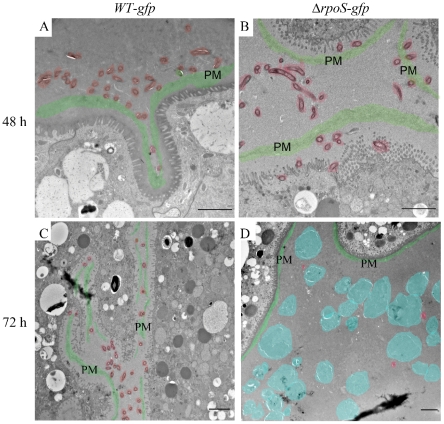

Similar articles
-
Two Distinct Mechanisms Govern RpoS-Mediated Repression of Tick-Phase Genes during Mammalian Host Adaptation by Borrelia burgdorferi, the Lyme Disease Spirochete.mBio. 2017 Aug 22;8(4):e01204-17. doi: 10.1128/mBio.01204-17. mBio. 2017. PMID: 28830947 Free PMC article.
-
BosR and PlzA reciprocally regulate RpoS function to sustain Borrelia burgdorferi in ticks and mammals.J Clin Invest. 2023 Mar 1;133(5):e166710. doi: 10.1172/JCI166710. J Clin Invest. 2023. PMID: 36649080 Free PMC article.
-
Analysis of the RpoS regulon in Borrelia burgdorferi in response to mammalian host signals provides insight into RpoS function during the enzootic cycle.Mol Microbiol. 2007 Sep;65(5):1193-217. doi: 10.1111/j.1365-2958.2007.05860.x. Epub 2007 Jul 23. Mol Microbiol. 2007. PMID: 17645733 Free PMC article.
-
Gene regulation in Borrelia burgdorferi.Annu Rev Microbiol. 2011;65:479-99. doi: 10.1146/annurev.micro.112408.134040. Annu Rev Microbiol. 2011. PMID: 21801026 Review.
-
Borrelia burgdorferi--traveling incognito?Microbes Infect. 2006 Apr;8(5):1390-9. doi: 10.1016/j.micinf.2005.12.022. Epub 2006 Mar 24. Microbes Infect. 2006. PMID: 16698304 Review.
Cited by
-
The Stringent Response-Regulated sRNA Transcriptome of Borrelia burgdorferi.Front Cell Infect Microbiol. 2018 Jul 5;8:231. doi: 10.3389/fcimb.2018.00231. eCollection 2018. Front Cell Infect Microbiol. 2018. PMID: 30027068 Free PMC article.
-
Metamorphoses of Lyme disease spirochetes: phenomenon of Borrelia persisters.Parasit Vectors. 2019 May 16;12(1):237. doi: 10.1186/s13071-019-3495-7. Parasit Vectors. 2019. PMID: 31097026 Free PMC article. Review.
-
Lipoprotein succession in Borrelia burgdorferi: similar but distinct roles for OspC and VlsE at different stages of mammalian infection.Mol Microbiol. 2013 Jul;89(2):216-27. doi: 10.1111/mmi.12271. Epub 2013 Jun 7. Mol Microbiol. 2013. PMID: 23692497 Free PMC article.
-
DksA-dependent regulation of RpoS contributes to Borrelia burgdorferi tick-borne transmission and mammalian infectivity.PLoS Pathog. 2021 Feb 18;17(2):e1009072. doi: 10.1371/journal.ppat.1009072. eCollection 2021 Feb. PLoS Pathog. 2021. PMID: 33600418 Free PMC article.
-
The cyclic-di-GMP signaling pathway in the Lyme disease spirochete, Borrelia burgdorferi.Front Cell Infect Microbiol. 2014 May 1;4:56. doi: 10.3389/fcimb.2014.00056. eCollection 2014. Front Cell Infect Microbiol. 2014. PMID: 24822172 Free PMC article. Review.
References
-
- Gruber TM, Gross CA. Multiple sigma subunits and the partitioning of bacterial transcription space. Annu Rev Microbiol. 2003;57:441–466. - PubMed
-
- Hengge-Aronis R. Recent insights into the general stress response regulatory network in Escherichia coli. J Mol Microbiol Biotechnol. 2002;4:341–346. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
