

Efficient capture of infected neutrophils by dendritic cells in the skin inhibits the early anti-leishmania response
- PMID: 22359507
- PMCID: PMC3280984
- DOI: 10.1371/journal.ppat.1002536
Efficient capture of infected neutrophils by dendritic cells in the skin inhibits the early anti-leishmania response
Abstract
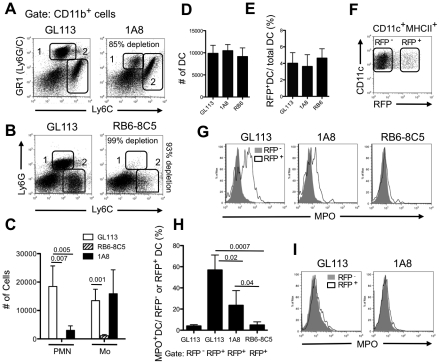
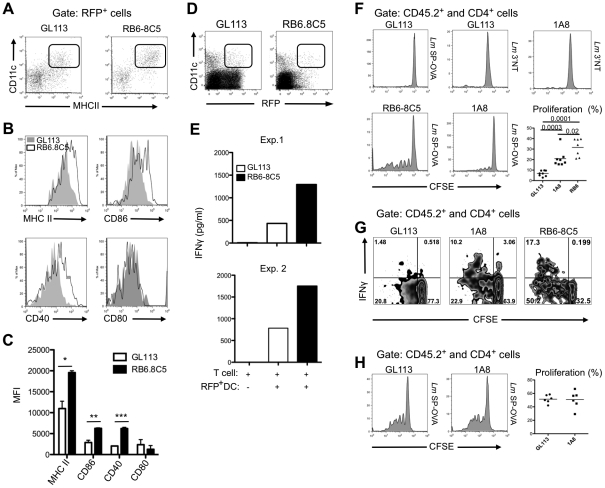
Neutrophils and dendritic cells (DCs) converge at localized sites of acute inflammation in the skin following pathogen deposition by the bites of arthropod vectors or by needle injection. Prior studies in mice have shown that neutrophils are the predominant recruited and infected cells during the earliest stage of Leishmania major infection in the skin, and that neutrophil depletion promotes host resistance to sand fly transmitted infection. How the massive influx of neutrophils aimed at wound repair and sterilization might modulate the function of DCs in the skin has not been previously addressed. The infected neutrophils recovered from the skin expressed elevated apoptotic markers compared to uninfected neutrophils, and were preferentially captured by dermal DCs when injected back into the mouse ear dermis. Following challenge with L. major directly, the majority of the infected DCs recovered from the skin at 24 hr stained positive for neutrophil markers, indicating that they acquired their parasites via uptake of infected neutrophils. When infected, dermal DCs were recovered from neutrophil depleted mice, their expression of activation markers was markedly enhanced, as was their capacity to present Leishmania antigens ex vivo. Neutrophil depletion also enhanced the priming of L. major specific CD4(+) T cells in vivo. The findings suggest that following their rapid uptake by neutrophils in the skin, L. major exploits the immunosuppressive effects associated with the apoptotic cell clearance function of DCs to inhibit the development of acquired resistance until the acute neutrophilic response is resolved.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
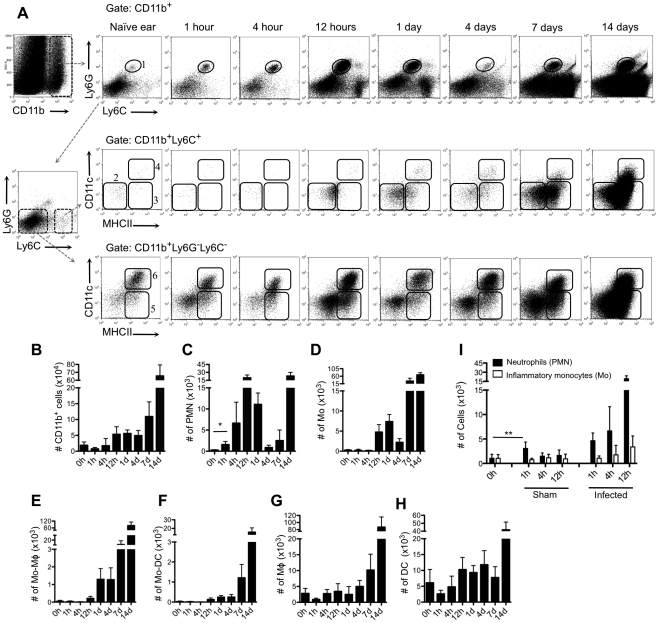
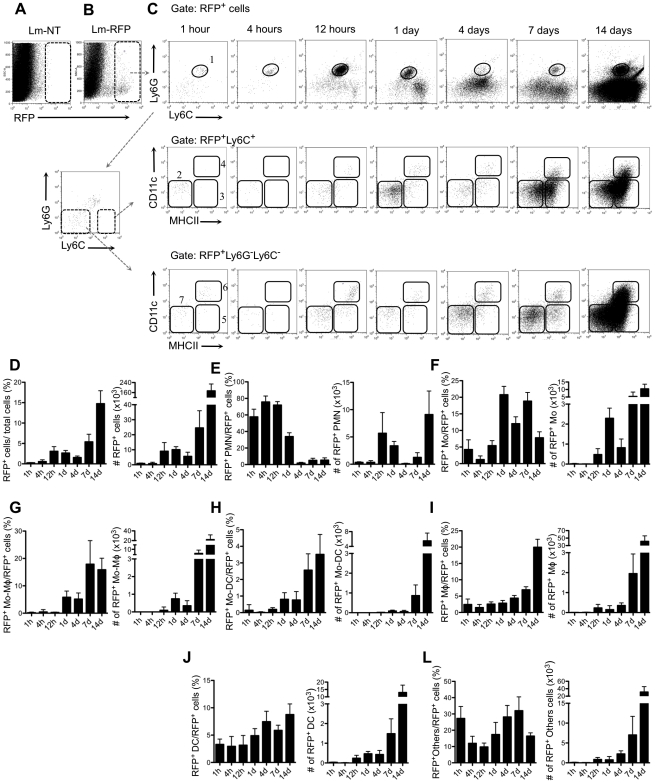
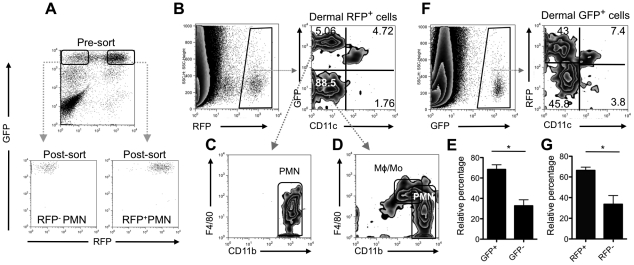
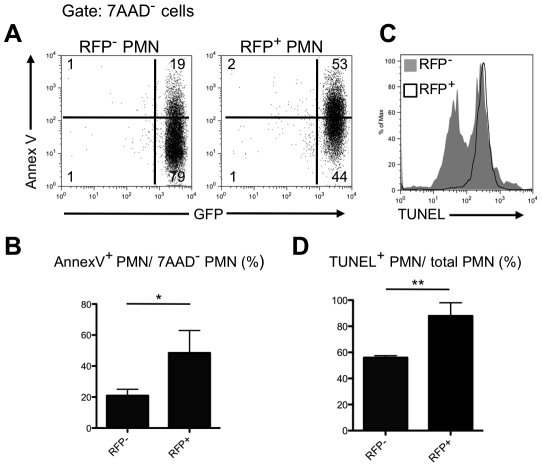
Similar articles
-
Apoptotic cell clearance of Leishmania major-infected neutrophils by dendritic cells inhibits CD8⁺ T-cell priming in vitro by Mer tyrosine kinase-dependent signaling.Cell Death Dis. 2015 Dec 10;6(12):e2018. doi: 10.1038/cddis.2015.351. Cell Death Dis. 2015. PMID: 26658192 Free PMC article.
-
In vivo imaging reveals an essential role for neutrophils in leishmaniasis transmitted by sand flies.Science. 2008 Aug 15;321(5891):970-4. doi: 10.1126/science.1159194. Science. 2008. PMID: 18703742 Free PMC article.
-
The role of dermis resident macrophages and their interaction with neutrophils in the early establishment of Leishmania major infection transmitted by sand fly bite.PLoS Pathog. 2020 Nov 2;16(11):e1008674. doi: 10.1371/journal.ppat.1008674. eCollection 2020 Nov. PLoS Pathog. 2020. PMID: 33137149 Free PMC article.
-
Dendritic Cells and Leishmania Infection: Adding Layers of Complexity to a Complex Disease.J Immunol Res. 2016;2016:3967436. doi: 10.1155/2016/3967436. Epub 2016 Jan 19. J Immunol Res. 2016. PMID: 26904694 Free PMC article. Review.
-
The Impact of Neutrophil Recruitment to the Skin on the Pathology Induced by Leishmania Infection.Front Immunol. 2021 Mar 1;12:649348. doi: 10.3389/fimmu.2021.649348. eCollection 2021. Front Immunol. 2021. PMID: 33732265 Free PMC article. Review.
Cited by
-
Determinants for the development of visceral leishmaniasis disease.PLoS Pathog. 2013 Jan;9(1):e1003053. doi: 10.1371/journal.ppat.1003053. Epub 2013 Jan 3. PLoS Pathog. 2013. PMID: 23300451 Free PMC article. Review.
-
Myeloid Cell Isolation from Mouse Skin and Draining Lymph Node Following Intradermal Immunization with Live Attenuated Plasmodium Sporozoites.J Vis Exp. 2016 May 18;(111):53796. doi: 10.3791/53796. J Vis Exp. 2016. PMID: 27286053 Free PMC article.
-
Clinical and immunological spectra of human cutaneous leishmaniasis in North Africa and French Guiana.Front Immunol. 2023 Jul 27;14:1134020. doi: 10.3389/fimmu.2023.1134020. eCollection 2023. Front Immunol. 2023. PMID: 37575260 Free PMC article. Review.
-
Leishmania Hijacks Myeloid Cells for Immune Escape.Front Microbiol. 2018 May 7;9:883. doi: 10.3389/fmicb.2018.00883. eCollection 2018. Front Microbiol. 2018. PMID: 29867798 Free PMC article. Review.
-
Apoptosis inhibition by intracellular bacteria and its consequence on host immunity.Curr Opin Immunol. 2019 Oct;60:103-110. doi: 10.1016/j.coi.2019.05.007. Epub 2019 Jun 19. Curr Opin Immunol. 2019. PMID: 31228759 Free PMC article. Review.
References
-
- Sacks D, Anderson C. Re-examination of the immunosuppressive mechanisms mediating non-cure of Leishmania infection in mice. Immunol Rev. 2004;201:225–238. - PubMed
-
- Belkaid Y, Mendez S, Lira R, Kadambi N, Milon G, et al. A natural model of Leishmania major infection reveals a prolonged “silent” phase of parasite amplification in the skin before the onset of lesion formation and immunity. J Immunol. 2000;165:969–977. - PubMed
-
- Laskay T, van Zandbergen G, Solbach W. Neutrophil granulocytes as host cells and transport vehicles for intracellular pathogens: apoptosis as infection-promoting factor. Immunobiology. 2008;213:183–191. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
