

A Co-Opted DEAD-Box RNA helicase enhances tombusvirus plus-strand synthesis
- PMID: 22359508
- PMCID: PMC3280988
- DOI: 10.1371/journal.ppat.1002537
A Co-Opted DEAD-Box RNA helicase enhances tombusvirus plus-strand synthesis
Abstract
Replication of plus-strand RNA viruses depends on recruited host factors that aid several critical steps during replication. In this paper, we show that an essential translation factor, Ded1p DEAD-box RNA helicase of yeast, directly affects replication of Tomato bushy stunt virus (TBSV). To separate the role of Ded1p in viral protein translation from its putative replication function, we utilized a cell-free TBSV replication assay and recombinant Ded1p. The in vitro data show that Ded1p plays a role in enhancing plus-strand synthesis by the viral replicase. We also find that Ded1p is a component of the tombusvirus replicase complex and Ded1p binds to the 3'-end of the viral minus-stranded RNA. The data obtained with wt and ATPase deficient Ded1p mutants support the model that Ded1p unwinds local structures at the 3'-end of the TBSV (-)RNA, rendering the RNA compatible for initiation of (+)-strand synthesis. Interestingly, we find that Ded1p and glyceraldehyde-3-phosphate dehydrogenase (GAPDH), which is another host factor for TBSV, play non-overlapping functions to enhance (+)-strand synthesis. Altogether, the two host factors enhance TBSV replication synergistically by interacting with the viral (-)RNA and the replication proteins. In addition, we have developed an in vitro assay for Flock house virus (FHV), a small RNA virus of insects, that also demonstrated positive effect on FHV replicase activity by the added Ded1p helicase. Thus, two small RNA viruses, which do not code for their own helicases, seems to recruit a host RNA helicase to aid their replication in infected cells.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
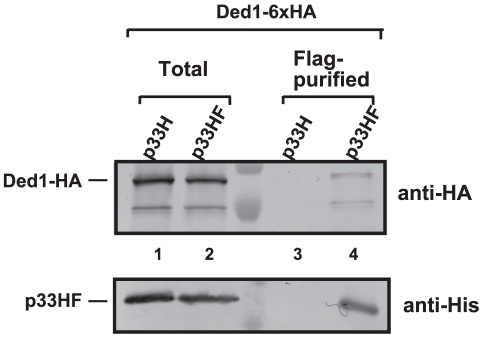


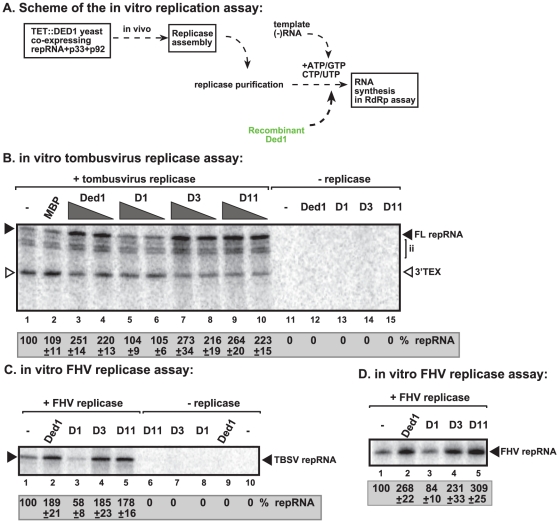



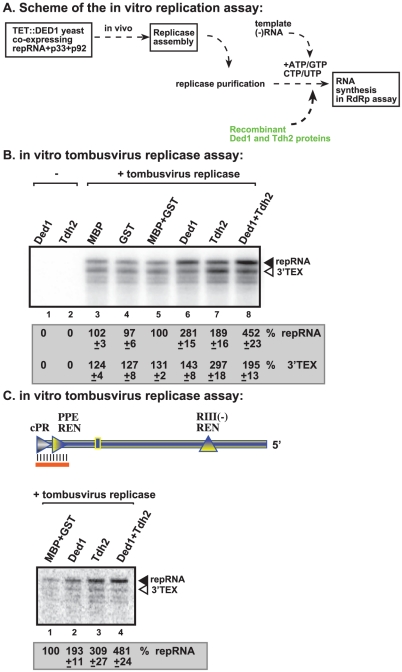

Similar articles
-
Similar roles for yeast Dbp2 and Arabidopsis RH20 DEAD-box RNA helicases to Ded1 helicase in tombusvirus plus-strand synthesis.Virology. 2012 Oct 25;432(2):470-84. doi: 10.1016/j.virol.2012.06.030. Epub 2012 Jul 24. Virology. 2012. PMID: 22832121
-
The expanding functions of cellular helicases: the tombusvirus RNA replication enhancer co-opts the plant eIF4AIII-like AtRH2 and the DDX5-like AtRH5 DEAD-box RNA helicases to promote viral asymmetric RNA replication.PLoS Pathog. 2014 Apr 17;10(4):e1004051. doi: 10.1371/journal.ppat.1004051. eCollection 2014 Apr. PLoS Pathog. 2014. PMID: 24743583 Free PMC article.
-
The proteasomal Rpn11 metalloprotease suppresses tombusvirus RNA recombination and promotes viral replication via facilitating assembly of the viral replicase complex.J Virol. 2015 Mar;89(5):2750-63. doi: 10.1128/JVI.02620-14. Epub 2014 Dec 24. J Virol. 2015. PMID: 25540361 Free PMC article.
-
Tombusvirus-Host Interactions: Co-Opted Evolutionarily Conserved Host Factors Take Center Court.Annu Rev Virol. 2016 Sep 29;3(1):491-515. doi: 10.1146/annurev-virology-110615-042312. Epub 2016 Aug 8. Annu Rev Virol. 2016. PMID: 27578441 Review.
-
Yeast as a model host to dissect functions of viral and host factors in tombusvirus replication.Virology. 2006 Jan 5;344(1):211-20. doi: 10.1016/j.virol.2005.09.017. Virology. 2006. PMID: 16364751 Review.
Cited by
-
Recruitment of Arabidopsis RNA Helicase AtRH9 to the Viral Replication Complex by Viral Replicase to Promote Turnip Mosaic Virus Replication.Sci Rep. 2016 Jul 26;6:30297. doi: 10.1038/srep30297. Sci Rep. 2016. PMID: 27456972 Free PMC article.
-
The centromeric histone CenH3 is recruited into the tombusvirus replication organelles.PLoS Pathog. 2022 Jun 29;18(6):e1010653. doi: 10.1371/journal.ppat.1010653. eCollection 2022 Jun. PLoS Pathog. 2022. PMID: 35767596 Free PMC article.
-
Blocking tombusvirus replication through the antiviral functions of DDX17-like RH30 DEAD-box helicase.PLoS Pathog. 2019 May 28;15(5):e1007771. doi: 10.1371/journal.ppat.1007771. eCollection 2019 May. PLoS Pathog. 2019. PMID: 31136641 Free PMC article.
-
Characterization of a nodavirus replicase revealed a de novo initiation mechanism of RNA synthesis and terminal nucleotidyltransferase activity.J Biol Chem. 2013 Oct 25;288(43):30785-801. doi: 10.1074/jbc.M113.492728. Epub 2013 Sep 9. J Biol Chem. 2013. PMID: 24019510 Free PMC article.
-
Co-opting the fermentation pathway for tombusvirus replication: Compartmentalization of cellular metabolic pathways for rapid ATP generation.PLoS Pathog. 2019 Oct 24;15(10):e1008092. doi: 10.1371/journal.ppat.1008092. eCollection 2019 Oct. PLoS Pathog. 2019. PMID: 31648290 Free PMC article.
References
-
- Nagy PD. Yeast as a model host to explore plant virus-host interactions. Annu Rev Phytopathol. 2008;46:217–242. - PubMed
-
- den Boon JA, Ahlquist P. Organelle-like membrane compartmentalization of positive-strand RNA virus replication factories. Annu Rev Microbiol. 2010;64:241–256. - PubMed
-
- Nagy PD, Wang RY, Pogany J, Hafren A, Makinen K. Emerging picture of host chaperone and cyclophilin roles in RNA virus replication. Virology. 2011;411:374–382. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
