

Disruption of MEF2C signaling and loss of sarcomeric and mitochondrial integrity in cancer-induced skeletal muscle wasting
- PMID: 22361433
- PMCID: PMC3314175
- DOI: 10.18632/aging.100436
Disruption of MEF2C signaling and loss of sarcomeric and mitochondrial integrity in cancer-induced skeletal muscle wasting
Abstract
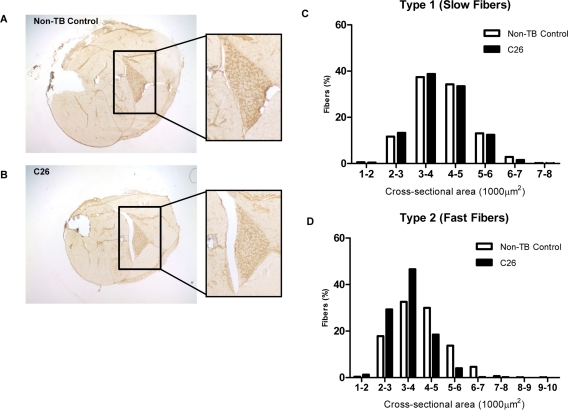
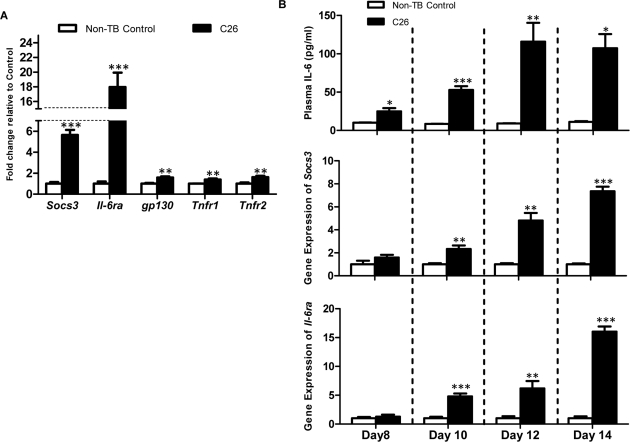
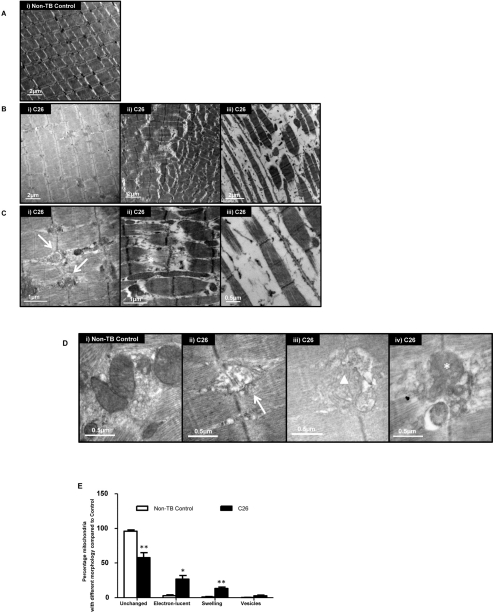
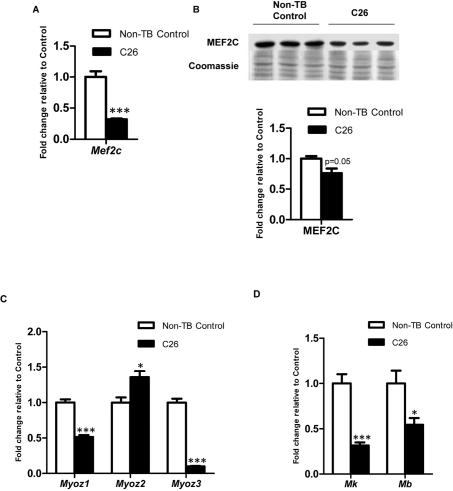
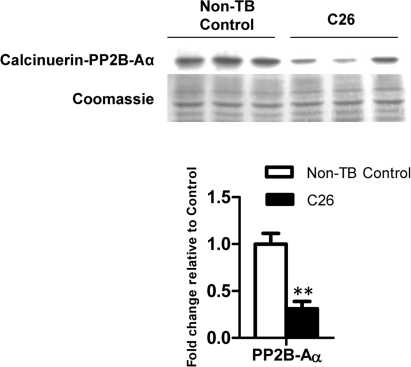
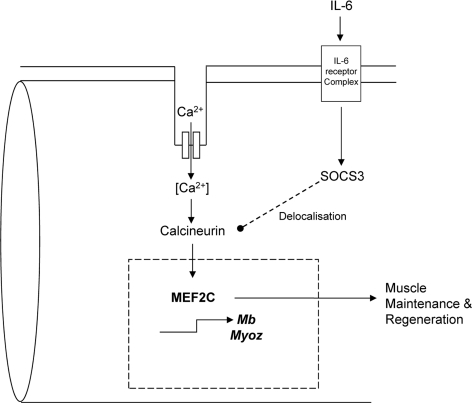
Cancer cachexia is a highly debilitating paraneoplastic disease observed in more than 50% of patients with advanced cancers and directly contributes to 20% of cancer deaths. Loss of skeletal muscle is a defining characteristic of patients with cancer cachexia and is associated with poor survival. The present study reveals the involvement of a myogenic transcription factor Myocyte Enhancer Factor (MEF) 2C in cancer-induced skeletal muscle wasting. Increased skeletal muscle mRNA expression of Suppressor of Cytokine Signaling (Socs) 3 and the IL-6 receptor indicative of active IL-6 signaling was seen in skeletal muscle of mice bearing the Colon 26 (C26) carcinoma. Loss of skeletal muscle structural integrity and distorted mitochondria were also observed using electron microscopy. Gene and protein expression of MEF2C was significantly downregulated in skeletal muscle from C26-bearing mice. MEF2C gene targets myozenin and myoglobin as well as myokinase were also altered during cachexia, suggesting dysregulated oxygen transport capacity and ATP regeneration in addition to distorted structural integrity. In addition, reduced expression of calcineurin was observed which suggested a potential pathway of MEF2C dysregulation. Together, these effects may limit sarcomeric contractile ability and also predispose skeletal muscle to structural instability; associated with muscle wasting and fatigue in cachexia.
Conflict of interest statement
The authors of this manuscript have no conflict of interest to declare.
Figures
References
-
- Tisdale MJ. Molecular pathways leading to cancer cachexia. Physiology (Bethesda) 2005;20:340–348. - PubMed
-
- Evans WJ, Morley JE, Argiles J, Bales C, Baracos V, Guttridge D, Jatoi A, Kalantar-Zadeh K, Lochs H, Mantovani G, Marks D, Mitch WE, Muscaritoli M, Najand A, Ponikowski P, Rossi Fanelli F, et al. Cachexia: a new definition. Clin Nutr. 2008;27:793–799. - PubMed
-
- Zhou X, Wang JL, Lu J, Song Y, Kwak KS, Jiao Q, Rosenfeld R, Chen Q, Boone T, Simonet WS, Lacey DL, Goldberg AL, Han HQ. Reversal of cancer cachexia and muscle wasting by ActRIIB antagonism leads to prolonged survival. Cell. 2010;142:531–543. - PubMed
-
- Guttridge DC, Mayo MW, Madrid LV, Wang CY, Baldwin AS., Jr. NF-kappaB-induced loss of MyoD messenger RNA: possible role in muscle decay and cachexia. Science. 2000;289:2363–2366. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
