

4,6-α-Glucanotransferase activity occurs more widespread in Lactobacillus strains and constitutes a separate GH70 subfamily
- PMID: 22361861
- PMCID: PMC3536977
- DOI: 10.1007/s00253-012-3943-1
4,6-α-Glucanotransferase activity occurs more widespread in Lactobacillus strains and constitutes a separate GH70 subfamily
Abstract
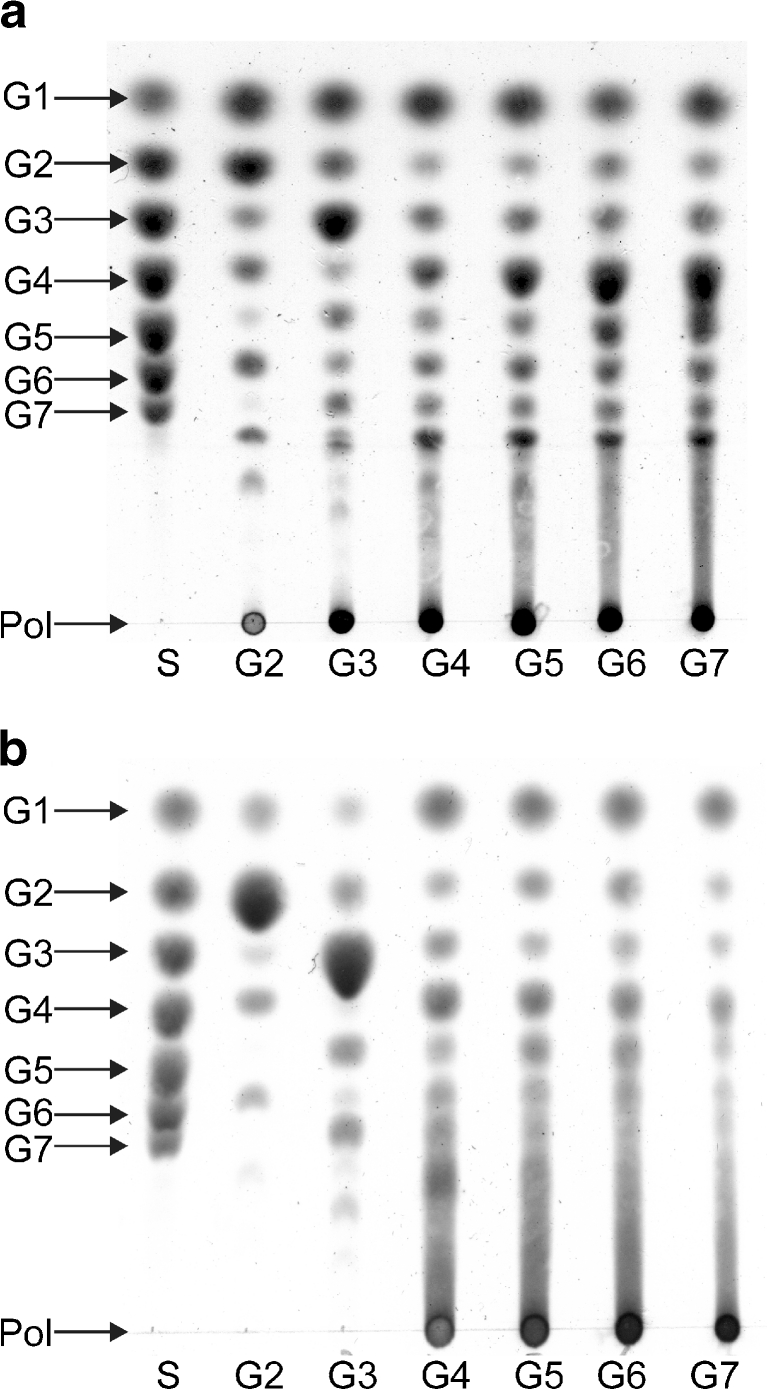
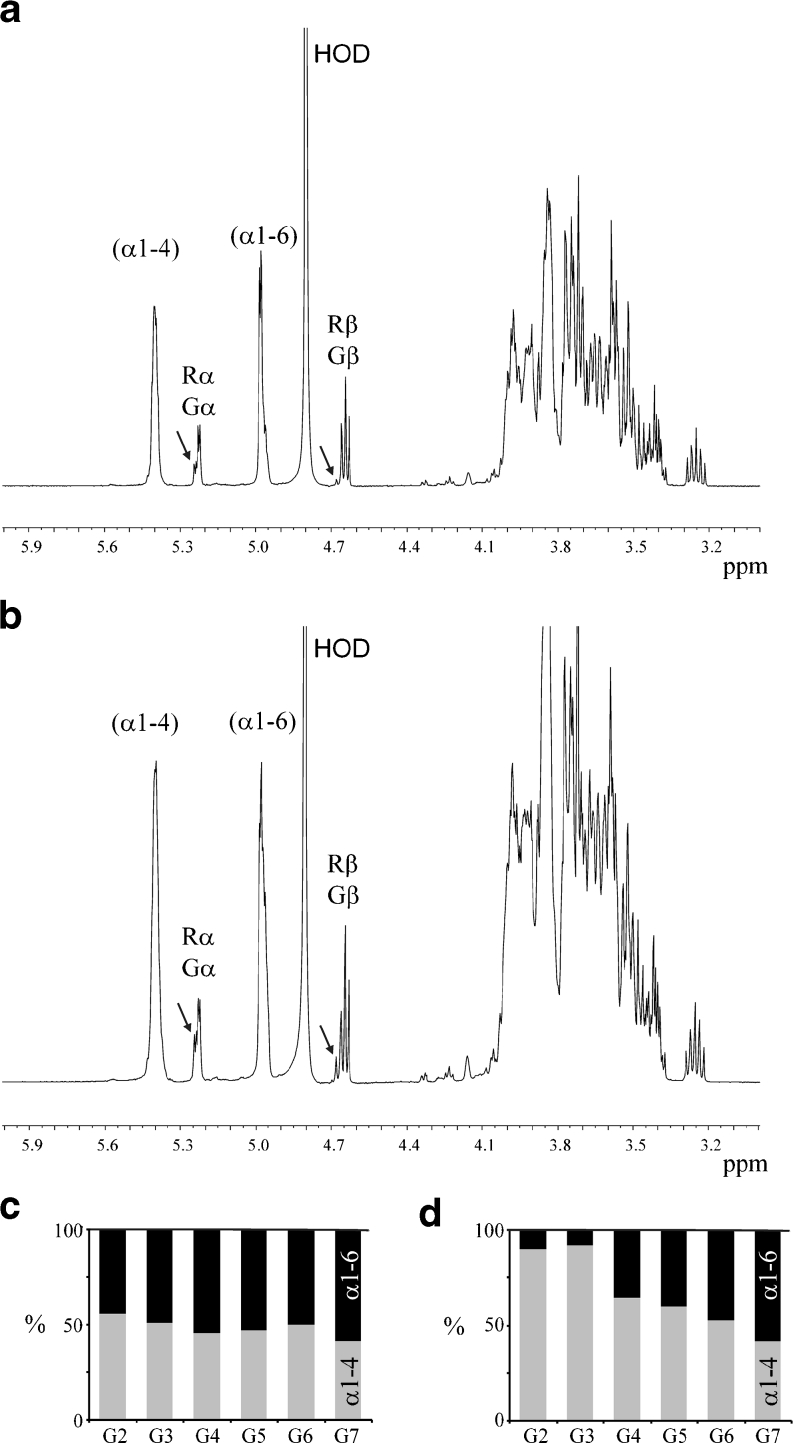
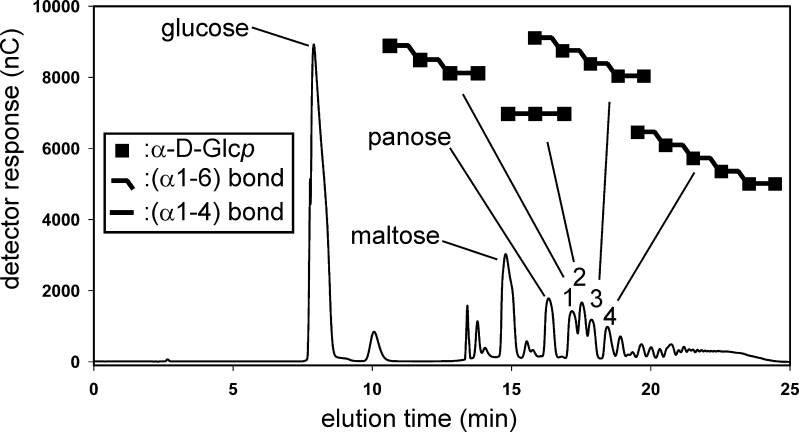
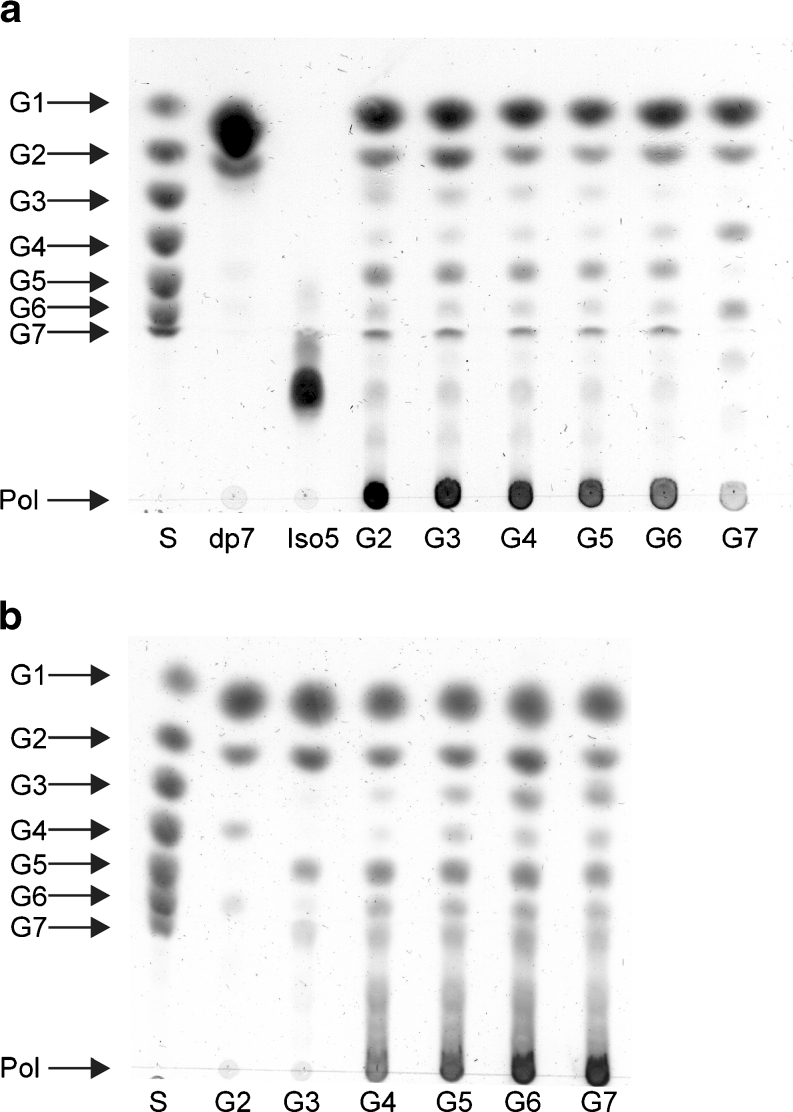
Family 70 glycoside hydrolase glucansucrase enzymes exclusively occur in lactic acid bacteria and synthesize a wide range of α-D-glucan (abbreviated as α-glucan) oligo- and polysaccharides. Of the 47 characterized GH70 enzymes, 46 use sucrose as glucose donor. A single GH70 enzyme was recently found to be inactive with sucrose and to utilize maltooligosaccharides [(1→4)-α-D-glucooligosaccharides] as glucose donor substrates for α-glucan synthesis, acting as a 4,6-α-glucanotransferase (4,6-αGT) enzyme. Here, we report the characterization of two further GH70 4,6-αGT enzymes, i.e., from Lactobacillus reuteri strains DSM 20016 and ML1, which use maltooligosaccharides as glucose donor. Both enzymes cleave α1→4 glycosidic linkages and add the released glucose moieties one by one to the non-reducing end of growing linear α-glucan chains via α1→6 glycosidic linkages (α1→4 to α1→6 transfer activity). In this way, they convert pure maltooligosaccharide substrates into linear α-glucan product mixtures with about 50% α1→6 glycosidic bonds (isomalto/maltooligosaccharides). These new α-glucan products may provide an exciting type of carbohydrate for the food industry. The results show that 4,6-αGTs occur more widespread in family GH70 and can be considered as a GH70 subfamily. Sequence analysis allowed identification of amino acid residues in acceptor substrate binding subsites +1 and +2, differing between GH70 GTF and 4,6-αGT enzymes.
Figures


References
-
- Albenne C, Skov LK, Mirza O, Gajhede M, Feller G, D'Amico S, André G, Potocki-Véronèse G, van der Veen BA, Monsan P, Remaud-Siméon M. Molecular basis of the amylose-like polymer formation catalyzed by Neisseria polysaccharea amylosucrase. J Biol Chem. 2004;279:726–734. doi: 10.1074/jbc.M309891200. - DOI - PubMed
-
- Brison Y, Pijning T, Malbert Y, Fabre E, Mourey L, Morel S, Potocki-Véronèse G, Monsan P, Tranier S, Remaud-Siméon M, Dijkstra BW (2012) Functional and structural characterization of an α-(1→2) branching sucrase derived from DSR-E glucansucrase. J Biol Chem. doi:10.1074/jbc.M111.305078 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
