

Junctions between i-motif tetramers in supramolecular structures
- PMID: 22362739
- PMCID: PMC3367196
- DOI: 10.1093/nar/gks161
Junctions between i-motif tetramers in supramolecular structures
Abstract
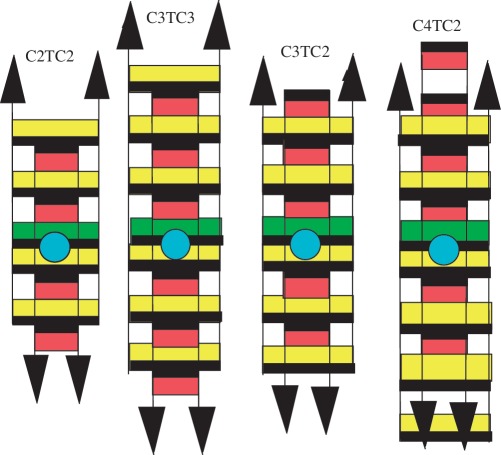
The symmetry of i-motif tetramers gives to cytidine-rich oligonucleotides the capacity to associate into supramolecular structures (sms). In order to determine how the tetramers are linked together in such structures, we have measured by gel filtration chromatography and NMR the formation and dissociation kinetics of sms built by oligonucleotides containing two short C stretches separated by a non-cytidine-base. We show that a stretch of only two cytidines either at the 3'- or 5'-end is long enough to link the tetramers into sms. The analysis of the properties of sms formed by oligonucleotides differing by the length of the oligo-C stretches, the sequence orientation and the nature of the non-C base provides a model of the junction connecting the tetramers in sms.
Figures








References
-
- Langridge R, Rich A. Molecular structure of helical polycytidylic acid. Nature. 1963;198:725–728. - PubMed
-
- Gehring K, Leroy JL, Guéron M. A tetrameric DNA structure with protonated cytidine·cytidine base pairs. Nature. 1993;363:561–565. - PubMed
-
- Guéron M, Leroy JL. The i-motif in nucleic acids. Curr. Opin. Struct. Biol. 2000;10:326–331. - PubMed
-
- Yan H, Park SH, Finkelstein G, Reif JH, LaBean TN. DNA-templated self-assembly of protein arrays and highly conductive nanowires. Science. 2003;301:1882–1884. - PubMed
