

Role of the neuronal K-Cl co-transporter KCC2 in inhibitory and excitatory neurotransmission
- PMID: 22363264
- PMCID: PMC3282916
- DOI: 10.3389/fncel.2012.00005
Role of the neuronal K-Cl co-transporter KCC2 in inhibitory and excitatory neurotransmission
Abstract
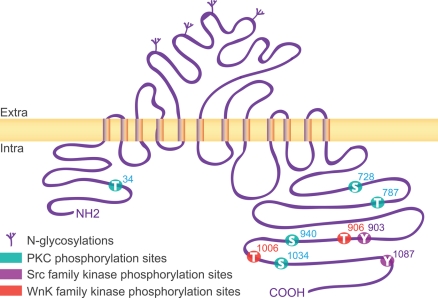
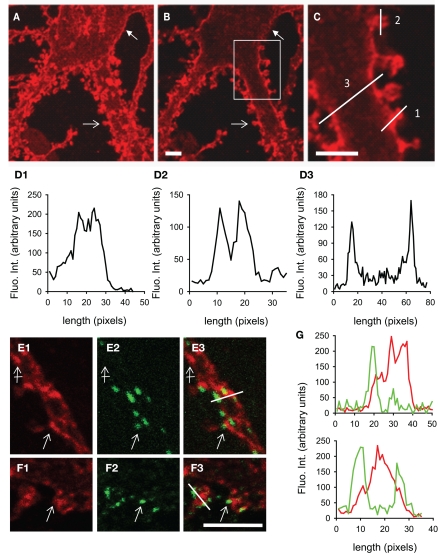
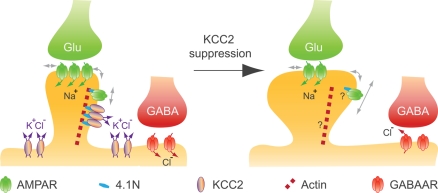
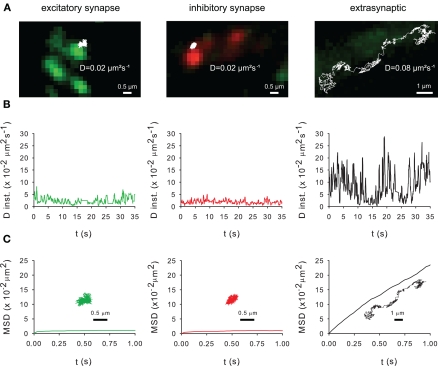
The K-Cl co-transporter KCC2 plays multiple roles in the physiology of central neurons and alterations of its function and/or expression are associated with several neurological conditions. By regulating intraneuronal chloride homeostasis, KCC2 strongly influences the efficacy and polarity of the chloride-permeable γ-aminobutyric acid (GABA) type A and glycine receptor (GlyR) mediated synaptic transmission. This appears particularly critical for the development of neuronal circuits as well as for the dynamic control of GABA and glycine signaling in mature networks. The activity of the transporter is also associated with transmembrane water fluxes which compensate solute fluxes associated with synaptic activity. Finally, KCC2 interaction with the actin cytoskeleton appears critical both for dendritic spine morphogenesis and the maintenance of glutamatergic synapses. In light of the pivotal role of KCC2 in the maturation and function of central synapses, it is of particular importance to understand the cellular and molecular mechanisms underlying its regulation. These include development and activity-dependent modifications both at the transcriptional and post-translational levels. We emphasize the importance of post-translational mechanisms such as phosphorylation and dephosphorylation, oligomerization, cell surface stability, clustering and membrane diffusion for the rapid and dynamic regulation of KCC2 function.
Keywords: KCC2; excitatory and inhibitory synapses; neuronal activity; post-translational regulation.
Figures

References
-
- Aguado F., Carmona M. A., Pozas E., Aguilo A., Martinez-Guijarro F. J., Alcantara S., Borrell V., Yuste R., Ibanez C. F., Soriano E. (2003). BDNF regulates spontaneous correlated activity at early developmental stages by increasing synaptogenesis and expression of the K+/Cl− co-transporter KCC2. Development 130, 1267–1280 10.1242/dev.00351 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources