

Possible role of acetylcholine in regulating spatial novelty effects on theta rhythm and grid cells
- PMID: 22363266
- PMCID: PMC3282552
- DOI: 10.3389/fncir.2012.00005
Possible role of acetylcholine in regulating spatial novelty effects on theta rhythm and grid cells
Abstract
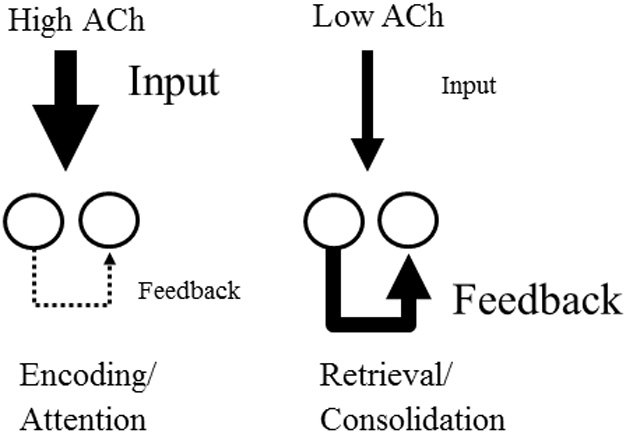
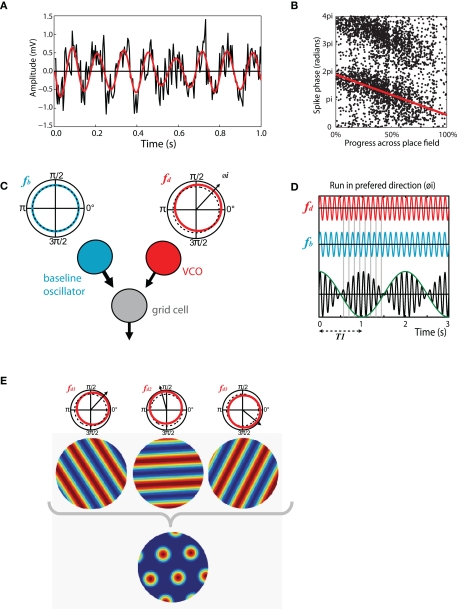
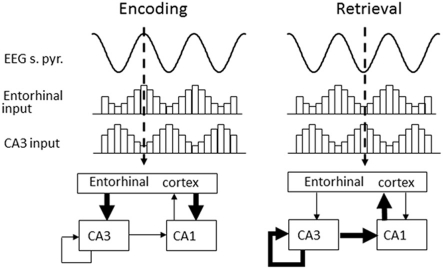
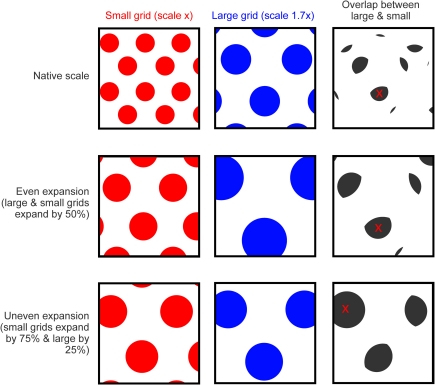
Existing pharmacological and lesion data indicate that acetylcholine plays an important role in memory formation. For example, increased levels of acetylcholine in the hippocampal formation are known to be associated with successful encoding while disruption of the cholinergic system leads to impairments on a range of mnemonic tasks. However, cholinergic signaling from the medial septum also plays a central role in generating and pacing theta-band oscillations throughout the hippocampal formation. Recent experimental results suggest a potential link between these distinct phenomena. Environmental novelty, a condition associated with strong cholinergic drive, has been shown to induce an expansion in the firing pattern of entorhinal grid cells and a reduction in the frequency of theta measured from the LFP. Computational modeling suggests the spatial activity of grid cells is produced by interference between neuronal oscillators; scale being determined by theta-band oscillations impinging on entorhinal stellate cells, the frequency of which is modulated by acetylcholine. Here we propose that increased cholinergic signaling in response to environmental novelty triggers grid expansion by reducing the frequency of the oscillations. Furthermore, we argue that cholinergic induced grid expansion may enhance, or even induce, encoding by producing a mismatch between expanded grid cells and other spatial inputs to the hippocampus, such as boundary vector cells. Indeed, a further source of mismatch is likely to occur between grid cells of different native scales which may expand by different relative amounts.
Keywords: acetylcholine; grid cell; place cell; stellate cell; theta.
Figures
References
-
- Aigner T. G., Mishkin M. (1986). The effects of physostigmine and scopolamine on recognition memory in monkeys. Behav. Neurosci. 45, 81–87 - PubMed
-
- Alonso A., Klink R. (1993). Differential electroresponsiveness of stellate and pyramidal-like cells of medial entorhinal cortex layer II. J. Neurophysiol. 70, 128–143 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources