

Modeling activity and target-dependent developmental cell death of mouse retinal ganglion cells ex vivo
- PMID: 22363559
- PMCID: PMC3281910
- DOI: 10.1371/journal.pone.0031105
Modeling activity and target-dependent developmental cell death of mouse retinal ganglion cells ex vivo
Abstract
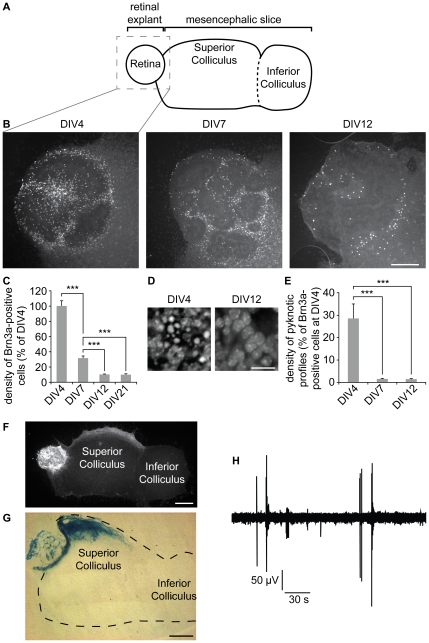
Programmed cell death is widespread during the development of the central nervous system and serves multiple purposes including the establishment of neural connections. In the mouse retina a substantial reduction of retinal ganglion cells (RGCs) occurs during the first postnatal week, coinciding with the formation of retinotopic maps in the superior colliculus (SC). We previously established a retino-collicular culture preparation which recapitulates the progressive topographic ordering of RGC projections during early post-natal life. Here, we questioned whether this model could also be suitable to examine the mechanisms underlying developmental cell death of RGCs. Brn3a was used as a marker of the RGCs. A developmental decline in the number of Brn3a-immunolabelled neurons was found in the retinal explant with a timing that paralleled that observed in vivo. In contrast, the density of photoreceptors or of starburst amacrine cells increased, mimicking the evolution of these cell populations in vivo. Blockade of neural activity with tetrodotoxin increased the number of surviving Brn3a-labelled neurons in the retinal explant, as did the increase in target availability when one retinal explant was confronted with 2 or 4 collicular slices. Thus, this ex vivo model reproduces the developmental reduction of RGCs and recapitulates its regulation by neural activity and target availability. It therefore offers a simple way to analyze developmental cell death in this classic system. Using this model, we show that ephrin-A signaling does not participate to the regulation of the Brn3a population size in the retina, indicating that eprhin-A-mediated elimination of exuberant projections does not involve developmental cell death.
Conflict of interest statement
Figures






References
-
- Buss RR, Sun W, Oppenheim RW. Adaptive roles of programmed cell death during nervous system development. Annu Rev Neurosci. 2006;29:1–35. - PubMed
-
- Vecino E, Hernández M, García M. Cell death in the developing vertebrate retina. Int J Dev Biol. 2004;48:965–974. - PubMed
-
- Young RW. Cell death during differentiation of the retina in the mouse. J Comp Neurol. 1984;229:362–373. - PubMed
-
- Young RW. Cell proliferation during postnatal development of the retina in the mouse. Brain Res. 1985;353:229–239. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
