

Establishment of motor neuron-V3 interneuron progenitor domain boundary in ventral spinal cord requires Groucho-mediated transcriptional corepression
- PMID: 22363571
- PMCID: PMC3281934
- DOI: 10.1371/journal.pone.0031176
Establishment of motor neuron-V3 interneuron progenitor domain boundary in ventral spinal cord requires Groucho-mediated transcriptional corepression
Abstract
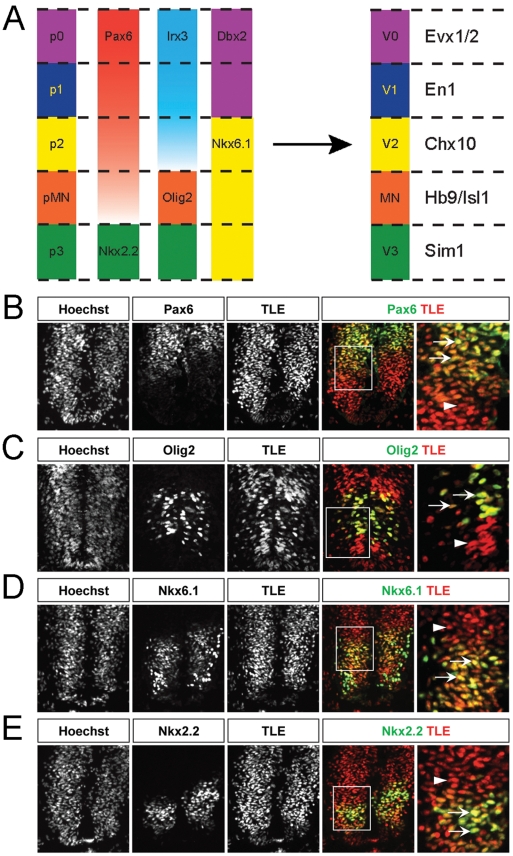
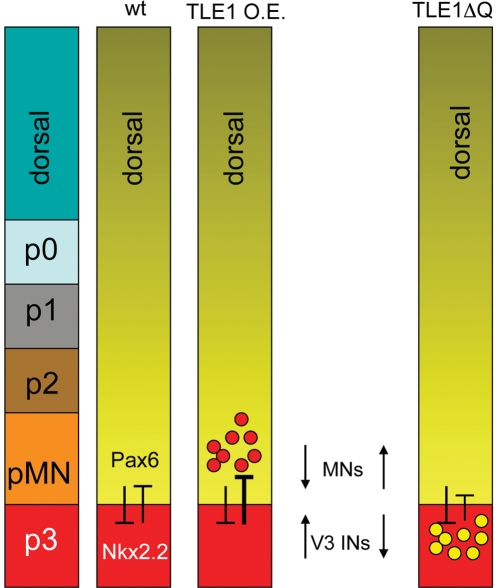
Background: Dorsoventral patterning of the developing spinal cord is important for the correct generation of spinal neuronal types. This process relies in part on cross-repressive interactions between specific transcription factors whose expression is regulated by Sonic hedgehog. Groucho/transducin-like Enhancer of split (TLE) proteins are transcriptional corepressors suggested to be recruited by at least certain Sonic hedgehog-controlled transcription factors to mediate the formation of spatially distinct progenitor domains within the ventral spinal cord. The aim of this study was to characterize the involvement of TLE in mechanisms regulating the establishment of the boundary between the most ventral spinal cord progenitor domains, termed pMN and p3. Because the pMN domain gives rise to somatic motor neurons while the p3 domain generates V3 interneurons, we also examined the involvement of TLE in the acquisition of these neuronal fates.
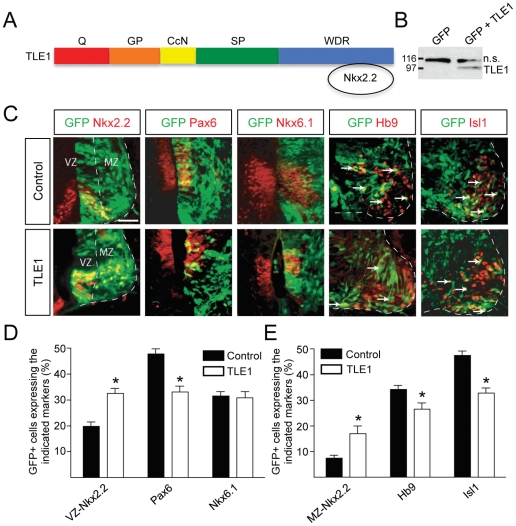
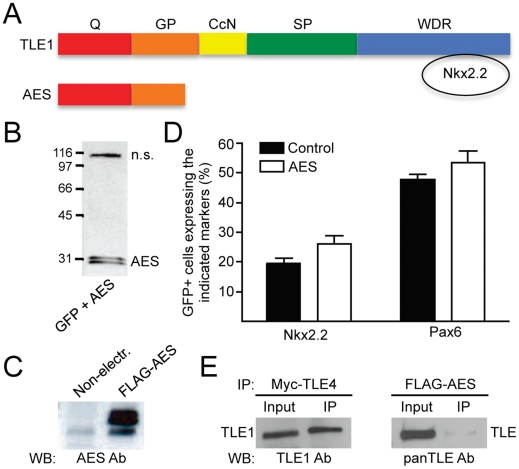
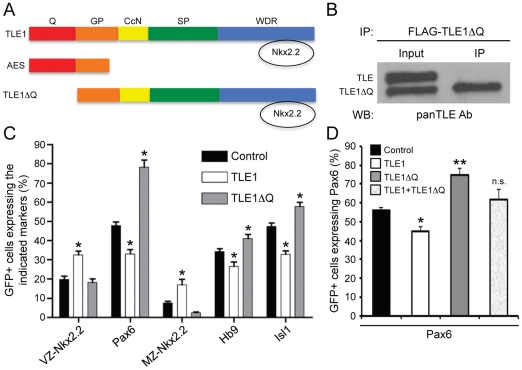
Methodology and principal findings: A combination of in vivo loss- and gain-of-function studies in the developing chick spinal cord was performed to characterize the role of TLE in ventral progenitor domain formation. It is shown here that TLE overexpression causes increased numbers of p3 progenitors and promotes the V3 interneuron fate while suppressing the motor neuron fate. Conversely, dominant-inhibition of TLE increases the numbers of pMN progenitors and postmitotic motor neurons.
Conclusion: Based on these results, we propose that TLE is important to promote the formation of the p3 domain and subsequent generation of V3 interneurons.
Conflict of interest statement
Figures
References
-
- Briscoe J, Ericson J. Specification of neuronal fates in the ventral neural tube. Curr Opin Neurobiol. 2001;11:43–49. - PubMed
-
- Zhuang B, Sockanathan S. Dorso-ventral patterning: a view from the top. Curr Opin Neurobiol. 2006;16:20–24. - PubMed
-
- Dessaud E, McMahon AP, Briscoe J. Pattern formation in the vertebrate neural tube: a sonic hedgehog morphogen-regulated transcriptional network. Development. 2008;135:2489–2503. - PubMed
-
- Lupo G, Harris WA, Lewis KE. Mechanisms of ventral patterning in the vertebrate nervous system. Nat Rev Neurosci. 2006;7:103–114. - PubMed
-
- Lu QR, Yuk D, Alberta JA, Zhu Z, Pawlitzky I, et al. Sonic hedgehog-regulated oligodendrocyte lineage genes encoding bHLH proteins in the mammalian central nervous system. Neuron. 2000;25:317–329. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
