

Preformed CD40L is stored in Th1, Th2, Th17, and T follicular helper cells as well as CD4+ 8- thymocytes and invariant NKT cells but not in Treg cells
- PMID: 22363608
- PMCID: PMC3283616
- DOI: 10.1371/journal.pone.0031296
Preformed CD40L is stored in Th1, Th2, Th17, and T follicular helper cells as well as CD4+ 8- thymocytes and invariant NKT cells but not in Treg cells
Abstract
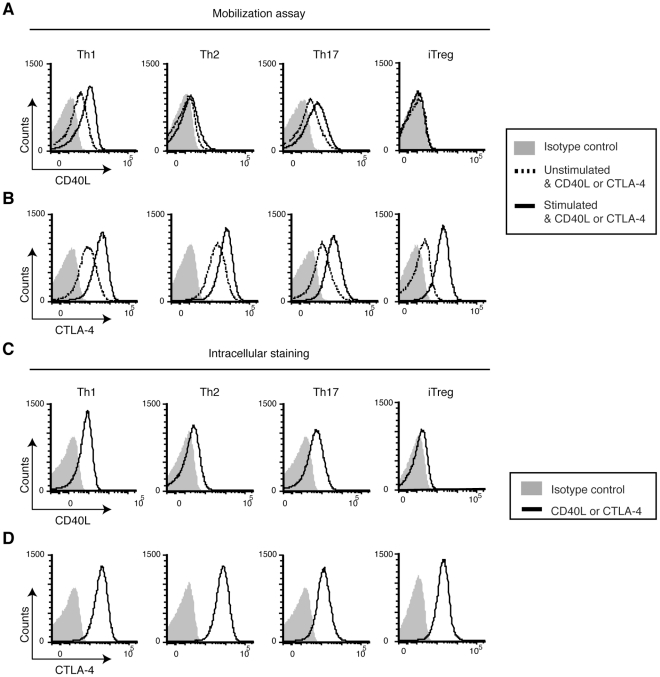
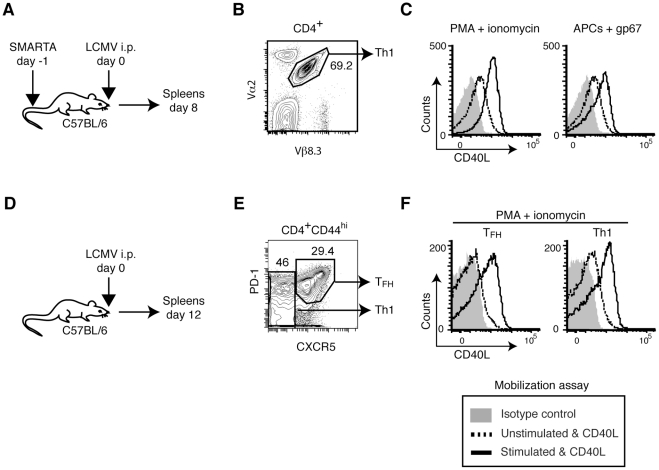
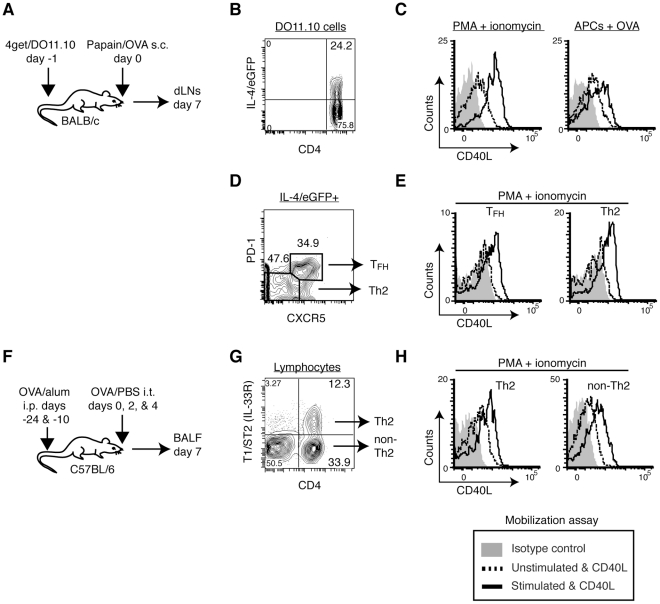
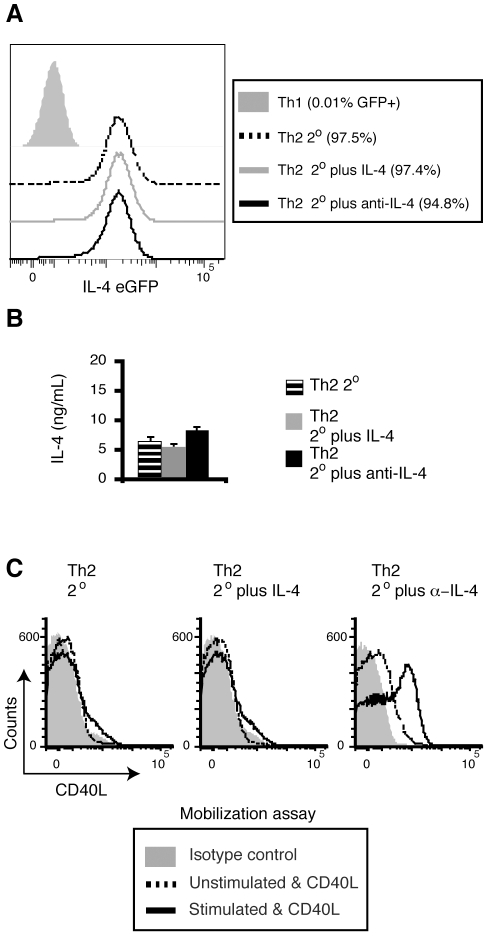
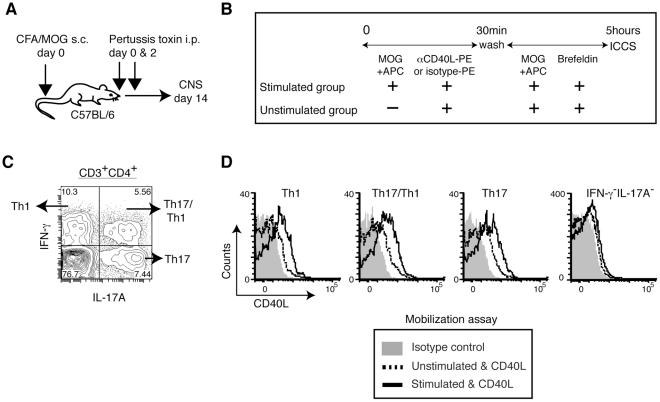
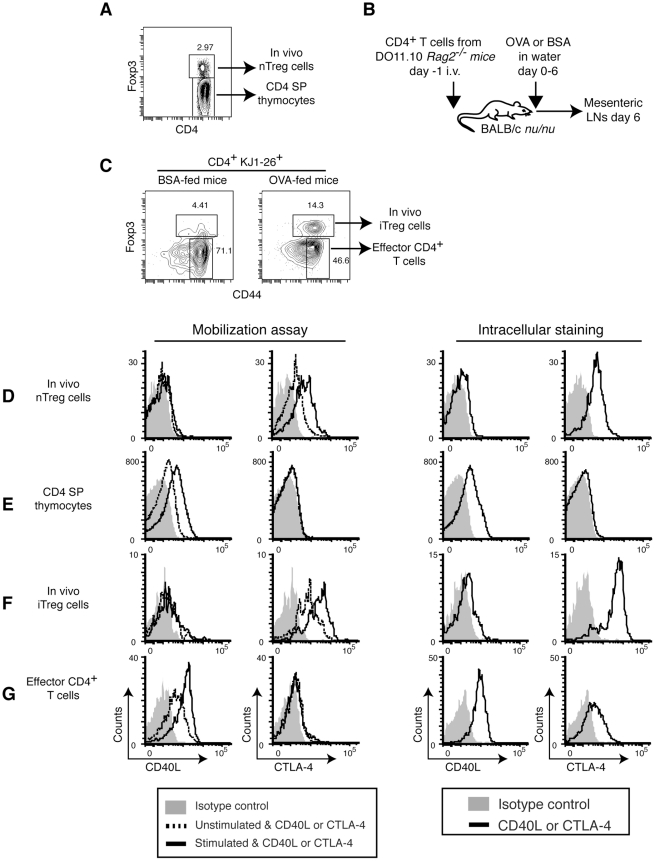
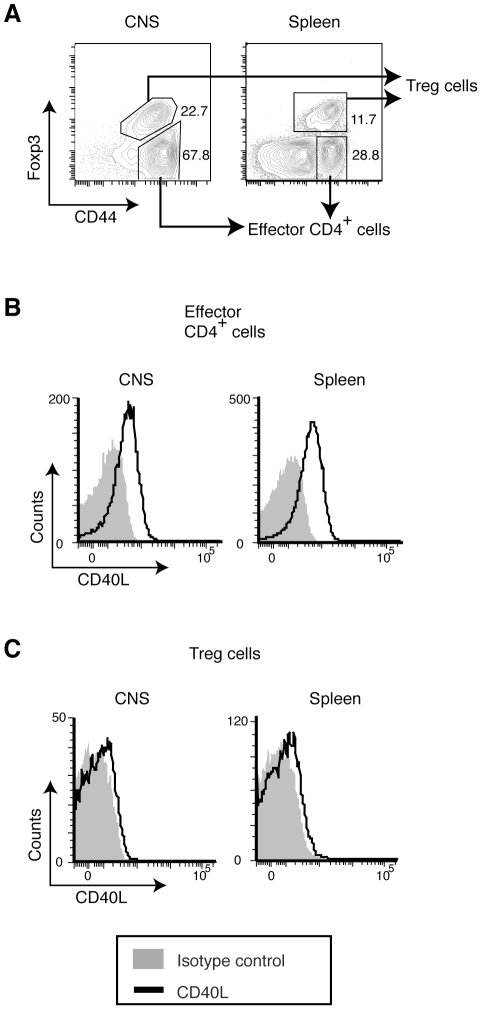
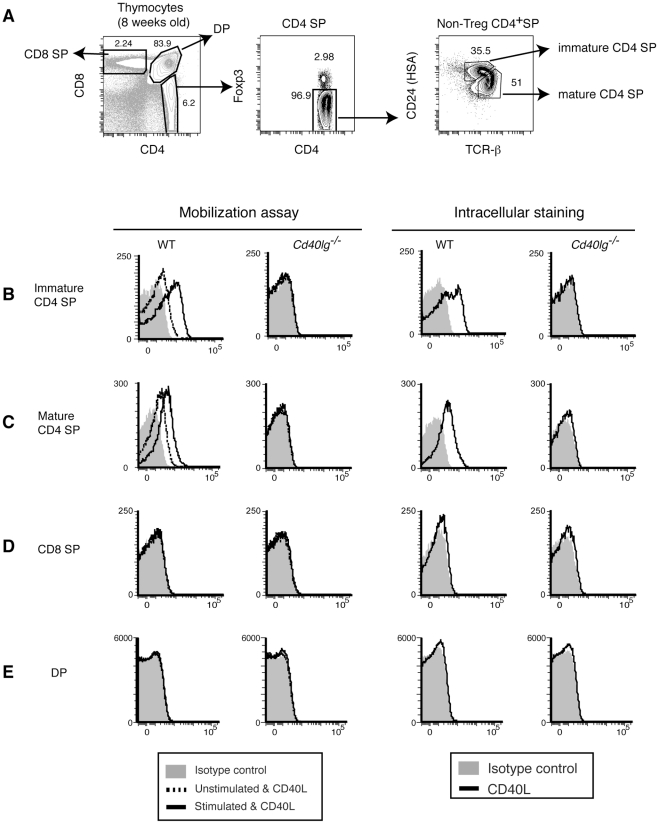
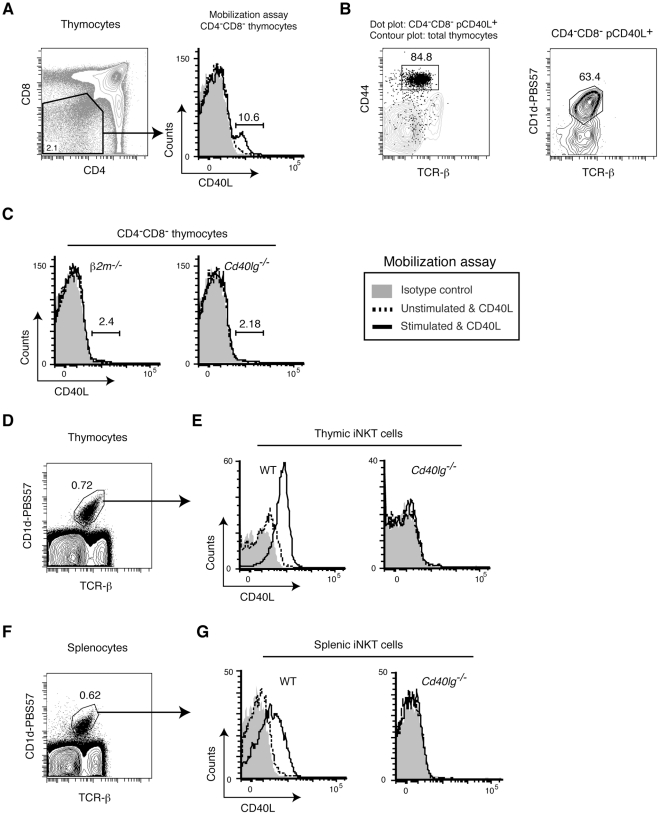
CD40L is essential for the development of adaptive immune responses. It is generally thought that CD40L expression in CD4(+) T cells is regulated transcriptionally and made from new mRNA following antigen recognition. However, imaging studies show that the majority of cognate interactions between effector CD4(+) T cells and APCs in vivo are too short to allow de novo CD40L synthesis. We previously showed that Th1 effector and memory cells store preformed CD40L (pCD40L) in lysosomal compartments and mobilize it onto the plasma membrane immediately after antigenic stimulation, suggesting that primed CD4(+) T cells may use pCD40L to activate APCs during brief encounters. Indeed, our recent study showed that pCD40L is sufficient to mediate selective activation of cognate B cells and trigger DC activation in vitro. In this study, we show that pCD40L is present in Th1 and follicular helper T cells developed during infection with lymphocytic choriomeningitis virus, Th2 cells in the airway of asthmatic mice, and Th17 cells from the CNS of animals with experimental autoimmune encephalitis (EAE). pCD40L is nearly absent in both natural and induced Treg cells, even in the presence of intense inflammation such as occurs in EAE. We also found pCD40L expression in CD4 single positive thymocytes and invariant NKT cells. Together, these results suggest that pCD40L may function in T cell development as well as an unexpectedly broad spectrum of innate and adaptive immune responses, while its expression in Treg cells is repressed to avoid compromising their suppressive activity.
Conflict of interest statement
Figures
References
-
- McHeyzer-Williams LJ, McHeyzer-Williams MG. Antigen-specific memory B cell development. Annu Rev Immunol. 2005;23:487–513. - PubMed
-
- Williams MA, Bevan MJ. Effector and memory CTL differentiation. Annu Rev Immunol. 2007;25:171–192. - PubMed
-
- Parker DC. T cell-dependent B cell activation. Annu Rev Immunol. 1993;11:331–360. - PubMed
-
- van Kooten C, Banchereau J. CD40-CD40 ligand. J Leukoc Biol. 2000;67:2–17. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI070934/AI/NIAID NIH HHS/United States
- HL061013/HL/NHLBI NIH HHS/United States
- HL071795/HL/NHLBI NIH HHS/United States
- R01 AI092080/AI/NIAID NIH HHS/United States
- R21 AI075064/AI/NIAID NIH HHS/United States
- K01 RR000163/RR/NCRR NIH HHS/United States
- RR000163/RR/NCRR NIH HHS/United States
- R01 HL071795/HL/NHLBI NIH HHS/United States
- R01 AI054458/AI/NIAID NIH HHS/United States
- P51 RR000163/RR/NCRR NIH HHS/United States
- R01 HL061013/HL/NHLBI NIH HHS/United States
- AI075064/AI/NIAID NIH HHS/United States
- R21 AI077032/AI/NIAID NIH HHS/United States
- AI054458/AI/NIAID NIH HHS/United States
- R0I AI050823/AI/NIAID NIH HHS/United States
- T32 AI078903/AI/NIAID NIH HHS/United States
- R01 AI050823/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
