

New technologies for 21st century plant science
- PMID: 22366161
- PMCID: PMC3315222
- DOI: 10.1105/tpc.111.093302
New technologies for 21st century plant science
Abstract
Plants are one of the most fascinating and important groups of organisms living on Earth. They serve as the conduit of energy into the biosphere, provide food, and shape our environment. If we want to make headway in understanding how these essential organisms function and build the foundation for a more sustainable future, then we need to apply the most advanced technologies available to the study of plant life. In 2009, a committee of the National Academy highlighted the "understanding of plant growth" as one of the big challenges for society and part of a new era which they termed "new biology." The aim of this article is to identify how new technologies can and will transform plant science to address the challenges of new biology. We assess where we stand today regarding current technologies, with an emphasis on molecular and imaging technologies, and we try to address questions about where we may go in the future and whether we can get an idea of what is at and beyond the horizon.
Figures

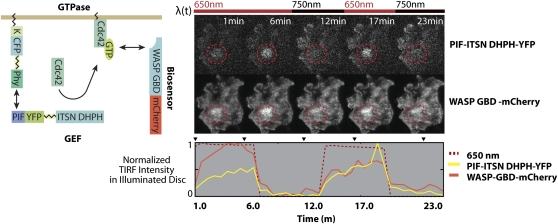





Similar articles
-
Big Data in Plant Science: Resources and Data Mining Tools for Plant Genomics and Proteomics.Methods Mol Biol. 2016;1415:533-47. doi: 10.1007/978-1-4939-3572-7_27. Methods Mol Biol. 2016. PMID: 27115651
-
Advances in plant genome sequencing.Plant J. 2012 Apr;70(1):177-90. doi: 10.1111/j.1365-313X.2012.04894.x. Plant J. 2012. PMID: 22449051 Review.
-
A decade of plant proteomics and mass spectrometry: translation of technical advancements to food security and safety issues.Mass Spectrom Rev. 2013 Sep-Oct;32(5):335-65. doi: 10.1002/mas.21365. Epub 2013 Jan 11. Mass Spectrom Rev. 2013. PMID: 23315723 Review.
-
Current state-of-art of sequencing technologies for plant genomics research.Brief Funct Genomics. 2012 Jan;11(1):3-11. doi: 10.1093/bfgp/elr045. Brief Funct Genomics. 2012. PMID: 22345601 Review.
-
Green systems biology - From single genomes, proteomes and metabolomes to ecosystems research and biotechnology.J Proteomics. 2011 Dec 10;75(1):284-305. doi: 10.1016/j.jprot.2011.07.010. Epub 2011 Jul 23. J Proteomics. 2011. PMID: 21802534 Review.
Cited by
-
Plant proteomics in India and Nepal: current status and challenges ahead.Physiol Mol Biol Plants. 2013 Oct;19(4):461-77. doi: 10.1007/s12298-013-0198-y. Physiol Mol Biol Plants. 2013. PMID: 24431515 Free PMC article. Review.
-
Dynamics and Endocytosis of Flot1 in Arabidopsis Require CPI1 Function.Int J Mol Sci. 2020 Feb 25;21(5):1552. doi: 10.3390/ijms21051552. Int J Mol Sci. 2020. PMID: 32106431 Free PMC article.
-
Editorial: Physiological, molecular and genetic perspectives of environmental stress response in plants.Front Plant Sci. 2023 Jun 16;14:1213762. doi: 10.3389/fpls.2023.1213762. eCollection 2023. Front Plant Sci. 2023. PMID: 37396640 Free PMC article. No abstract available.
-
Examination of Endogenous Peptides in Medicago truncatula Using Mass Spectrometry Imaging.J Proteome Res. 2016 Dec 2;15(12):4403-4411. doi: 10.1021/acs.jproteome.6b00471. Epub 2016 Oct 11. J Proteome Res. 2016. PMID: 27726374 Free PMC article.
-
Quinolizidine alkaloid biosynthesis: recent advances and future prospects.Front Plant Sci. 2012 Oct 26;3:239. doi: 10.3389/fpls.2012.00239. eCollection 2012. Front Plant Sci. 2012. PMID: 23112802 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
