

Dynamic denitrosylation via S-nitrosoglutathione reductase regulates cardiovascular function
- PMID: 22366318
- PMCID: PMC3306718
- DOI: 10.1073/pnas.1113319109
Dynamic denitrosylation via S-nitrosoglutathione reductase regulates cardiovascular function
Abstract
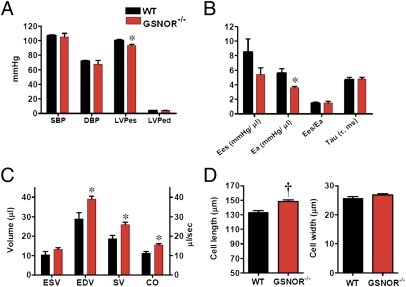
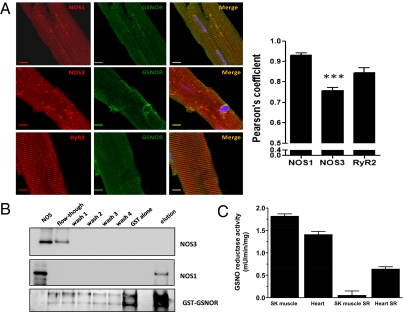
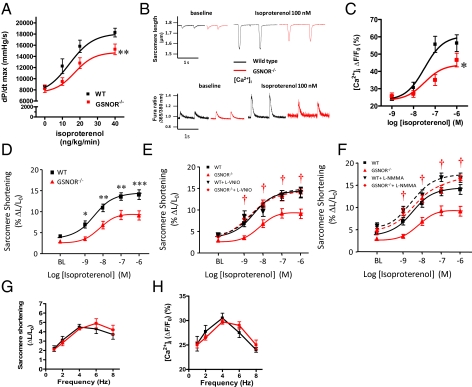
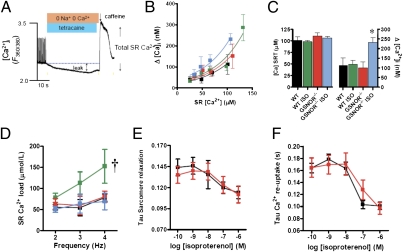
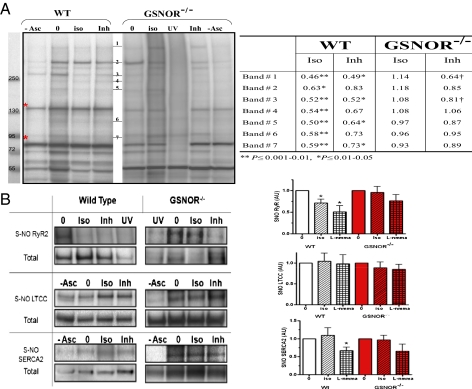
Although protein S-nitrosylation is increasingly recognized as mediating nitric oxide (NO) signaling, roles for protein denitrosylation in physiology remain unknown. Here, we show that S-nitrosoglutathione reductase (GSNOR), an enzyme that governs levels of S-nitrosylation by promoting protein denitrosylation, regulates both peripheral vascular tone and β-adrenergic agonist-stimulated cardiac contractility, previously ascribed exclusively to NO/cGMP. GSNOR-deficient mice exhibited reduced peripheral vascular tone and depressed β-adrenergic inotropic responses that were associated with impaired β-agonist-induced denitrosylation of cardiac ryanodine receptor 2 (RyR2), resulting in calcium leak. These results indicate that systemic hemodynamic responses (vascular tone and cardiac contractility), both under basal conditions and after adrenergic activation, are regulated through concerted actions of NO synthase/GSNOR and that aberrant denitrosylation impairs cardiovascular function. Our findings support the notion that dynamic S-nitrosylation/denitrosylation reactions are essential in cardiovascular regulation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Reduction of cardiomyocyte S-nitrosylation by S-nitrosoglutathione reductase protects against sepsis-induced myocardial depression.Am J Physiol Heart Circ Physiol. 2013 Apr 15;304(8):H1134-46. doi: 10.1152/ajpheart.00887.2012. Epub 2013 Feb 15. Am J Physiol Heart Circ Physiol. 2013. PMID: 23417863 Free PMC article.
-
Nitric Oxide Synthase 1 Modulates Basal and β-Adrenergic-Stimulated Contractility by Rapid and Reversible Redox-Dependent S-Nitrosylation of the Heart.PLoS One. 2016 Aug 16;11(8):e0160813. doi: 10.1371/journal.pone.0160813. eCollection 2016. PLoS One. 2016. PMID: 27529477 Free PMC article.
-
GSNOR Deficiency Enhances In Situ Skeletal Muscle Strength, Fatigue Resistance, and RyR1 S-Nitrosylation Without Impacting Mitochondrial Content and Activity.Antioxid Redox Signal. 2017 Feb 1;26(4):165-181. doi: 10.1089/ars.2015.6548. Epub 2016 Aug 19. Antioxid Redox Signal. 2017. PMID: 27412893 Free PMC article.
-
Nitrosative Stress and Human Disease: Therapeutic Potential of Denitrosylation.Int J Mol Sci. 2021 Sep 10;22(18):9794. doi: 10.3390/ijms22189794. Int J Mol Sci. 2021. PMID: 34575960 Free PMC article. Review.
-
Chronicles of a reductase: Biochemistry, genetics and physio-pathological role of GSNOR.Free Radic Biol Med. 2017 Sep;110:19-30. doi: 10.1016/j.freeradbiomed.2017.05.014. Epub 2017 May 19. Free Radic Biol Med. 2017. PMID: 28533171 Review.
Cited by
-
S-nitrosylation of CSF1 receptor increases the efficacy of CSF1R blockage against prostate cancer.Cell Death Dis. 2022 Oct 8;13(10):859. doi: 10.1038/s41419-022-05289-4. Cell Death Dis. 2022. PMID: 36209194 Free PMC article.
-
A postpartum enriched environment rescues impaired cognition and oxidative markers in aged mice with gestational inflammation.Brain Behav. 2022 Dec;12(12):e2817. doi: 10.1002/brb3.2817. Epub 2022 Nov 21. Brain Behav. 2022. PMID: 36409568 Free PMC article.
-
S-Nitrosylation of Sarcomeric Proteins Depresses Myofilament Ca2+)Sensitivity in Intact Cardiomyocytes.Antioxid Redox Signal. 2015 Nov 1;23(13):1017-34. doi: 10.1089/ars.2015.6275. Antioxid Redox Signal. 2015. PMID: 26421519 Free PMC article.
-
Oxidative, Reductive, and Nitrosative Stress Effects on Epigenetics and on Posttranslational Modification of Enzymes in Cardiometabolic Diseases.Oxid Med Cell Longev. 2020 Oct 30;2020:8819719. doi: 10.1155/2020/8819719. eCollection 2020. Oxid Med Cell Longev. 2020. PMID: 33204398 Free PMC article. Review.
-
Pharmacological Inhibition of S-Nitrosoglutathione Reductase Reduces Cardiac Damage Induced by Ischemia-Reperfusion.Antioxidants (Basel). 2021 Apr 2;10(4):555. doi: 10.3390/antiox10040555. Antioxidants (Basel). 2021. PMID: 33918310 Free PMC article.
References
-
- Murad F. Shattuck Lecture. Nitric oxide and cyclic GMP in cell signaling and drug development. N Engl J Med. 2006;355:2003–2011. - PubMed
-
- Gow AJ, et al. Basal and stimulated protein S-nitrosylation in multiple cell types and tissues. J Biol Chem. 2002;277:9637–9640. - PubMed
-
- González DR, et al. Differential role of S-nitrosylation and the NO-cGMP-PKG pathway in cardiac contractility. Nitric Oxide. 2008;18:157–167. - PubMed
-
- Liu L, et al. Essential roles of S-nitrosothiols in vascular homeostasis and endotoxic shock. Cell. 2004;116:617–628. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL091876/HL/NHLBI NIH HHS/United States
- HL059130/HL/NHLBI NIH HHS/United States
- R01 HL107110/HL/NHLBI NIH HHS/United States
- R01 HL084275/HL/NHLBI NIH HHS/United States
- HL095463/HL/NHLBI NIH HHS/United States
- R01 HL-094849/HL/NHLBI NIH HHS/United States
- HL91876/HL/NHLBI NIH HHS/United States
- R01 HL065455/HL/NHLBI NIH HHS/United States
- R01 HL-65455/HL/NHLBI NIH HHS/United States
- HL107110/HL/NHLBI NIH HHS/United States
- P01 HL-075443/HL/NHLBI NIH HHS/United States
- R01 HL110737/HL/NHLBI NIH HHS/United States
- R01 HL095463/HL/NHLBI NIH HHS/United States
- R01 HL094849/HL/NHLBI NIH HHS/United States
- HL084275/HL/NHLBI NIH HHS/United States
- R01 HL059130/HL/NHLBI NIH HHS/United States
- P01 HL075443/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
