

Impaired associative taste learning and abnormal brain activation in kinase-defective eEF2K mice
- PMID: 22366775
- PMCID: PMC3293518
- DOI: 10.1101/lm.023937.111
Impaired associative taste learning and abnormal brain activation in kinase-defective eEF2K mice
Abstract
Memory consolidation is defined temporally based on pharmacological interventions such as inhibitors of mRNA translation (molecular consolidation) or post-acquisition deactivation of specific brain regions (systems level consolidation). However, the relationship between molecular and systems consolidation are poorly understood. Molecular consolidation mechanisms involved in translation initiation and elongation have previously been studied in the cortex using taste-learning paradigms. For example, the levels of phosphorylation of eukaryotic elongation factor 2 (eEF2) were found to be correlated with taste learning in the gustatory cortex (GC), minutes following learning. In order to isolate the role of the eEF2 phosphorylation state at Thr-56 in both molecular and system consolidation, we analyzed cortical-dependent taste learning in eEF2K (the only known kinase for eEF2) ki mice, which exhibit reduced levels of eEF2 phosphorylation but normal levels of eEF2 and eEF2K. These mice exhibit clear attenuation of cortical-dependent associative, but not of incidental, taste learning. In order to gain a better understanding of the underlying mechanisms, we compared brain activity as measured by MEMRI (manganese-enhanced magnetic resonance imaging) between eEF2K ki mice and WT mice during conditioned taste aversion (CTA) learning and observed clear differences between the two but saw no differences under basal conditions. Our results demonstrate that adequate levels of phosphorylation of eEF2 are essential for cortical-dependent associative learning and suggest that malfunction of memory processing at the systems level underlies this associative memory impairment.
© 2012 Cold Spring Harbor Laboratory Press
Figures
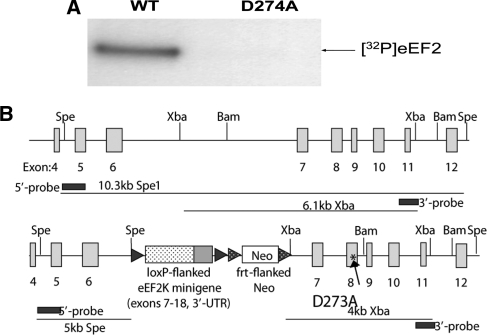
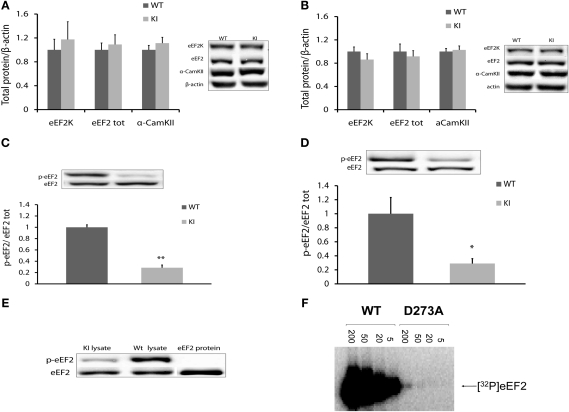
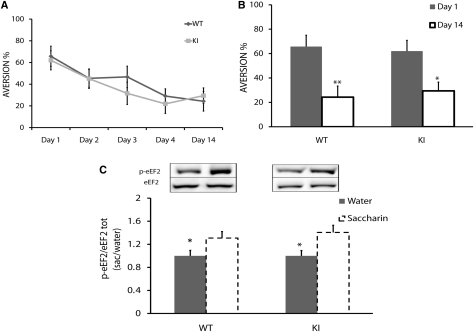
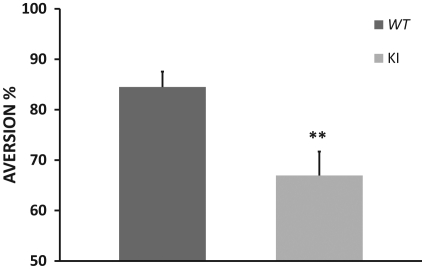
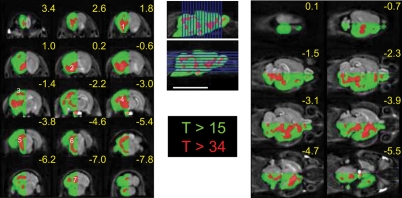
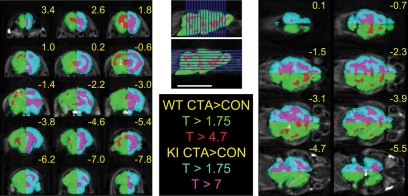
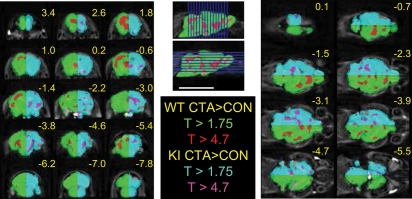
References
-
- Abel T, Lattal KM 2001. Molecular mechanisms of memory acquisition, consolidation, and retrieval. Curr Opin Neurobiol 11: 180–187 - PubMed
-
- Alberini CM 2005. Mechanisms of memory stabilization: Are consolidation and reconsolidation similar or distinct processes? Trends Neurosci 28: 51–56 - PubMed
-
- Aoki I, Naruse S, Tanaka C 2004. Manganese-enhanced magnetic resonance imaging (MEMRI) of brain activity and applications to early detection of brain ischemia. NMR Biomed 17: 569–580 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous