

Compound inheritance of a low-frequency regulatory SNP and a rare null mutation in exon-junction complex subunit RBM8A causes TAR syndrome
- PMID: 22366785
- PMCID: PMC3428915
- DOI: 10.1038/ng.1083
Compound inheritance of a low-frequency regulatory SNP and a rare null mutation in exon-junction complex subunit RBM8A causes TAR syndrome
Abstract
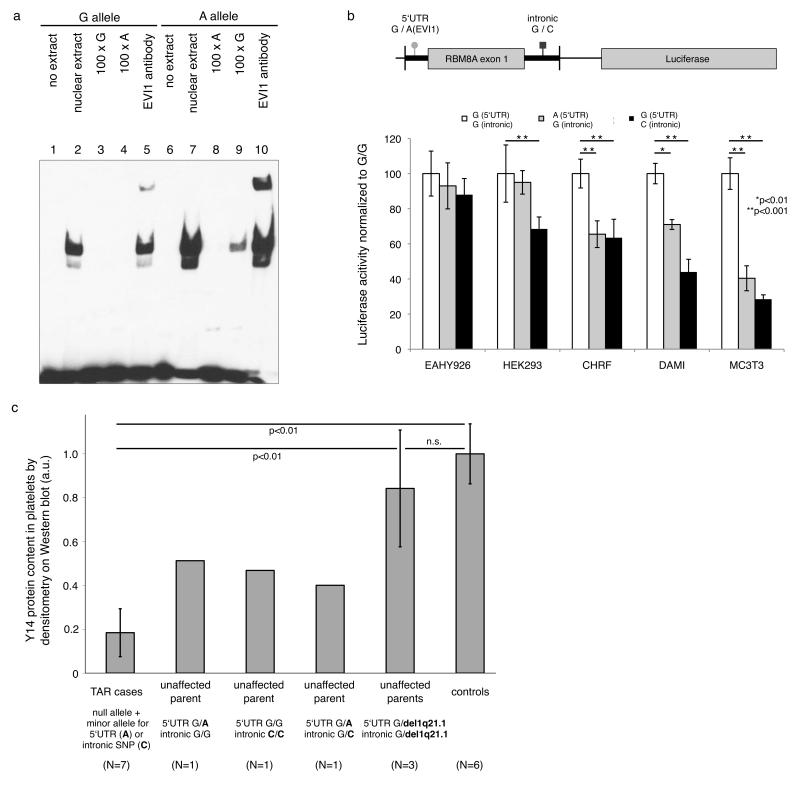
The exon-junction complex (EJC) performs essential RNA processing tasks. Here, we describe the first human disorder, thrombocytopenia with absent radii (TAR), caused by deficiency in one of the four EJC subunits. Compound inheritance of a rare null allele and one of two low-frequency SNPs in the regulatory regions of RBM8A, encoding the Y14 subunit of EJC, causes TAR. We found that this inheritance mechanism explained 53 of 55 cases (P < 5 × 10(-228)) of the rare congenital malformation syndrome. Of the 53 cases with this inheritance pattern, 51 carried a submicroscopic deletion of 1q21.1 that has previously been associated with TAR, and two carried a truncation or frameshift null mutation in RBM8A. We show that the two regulatory SNPs result in diminished RBM8A transcription in vitro and that Y14 expression is reduced in platelets from individuals with TAR. Our data implicate Y14 insufficiency and, presumably, an EJC defect as the cause of TAR syndrome.
Figures

Comment in
-
Deficiency of the Y14 protein is a critical factor underlying the etiology of thrombocytopenia with absent radii syndrome.Clin Genet. 2012 Jul;82(1):29-30. doi: 10.1111/j.1399-0004.2012.01888.x. Epub 2012 Apr 25. Clin Genet. 2012. PMID: 22486491 No abstract available.
References
-
- Kim VN, Kataoka N, Dreyfuss G. Role of the Nonsense-Mediated Decay Factor hUpf3 in the Splicing-Dependent Exon-Exon Junction Complex. Science. 2001;293:1832–1836. - PubMed
-
- Lykke-Andersen J, Shu M-D, Steitz JA. Communication of the Position of Exon-Exon Junctions to the mRNA Surveillance Machinery by the Protein RNPS1. Science. 2001;293:1836–1839. - PubMed
-
- Palacios IM, Gatfield D, St Johnston D, Izaurralde E. An eIF4AIII-containing complex required for mRNA localization and nonsense-mediated mRNA decay. Nature. 2004;427:753–757. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
- RP-PG-0310-1002/DH_/Department of Health/United Kingdom
- FS/09/039/27788/BHF_/British Heart Foundation/United Kingdom
- RG/08/014/24067/BHF_/British Heart Foundation/United Kingdom
- FS/09/039/BHF_/British Heart Foundation/United Kingdom
- WT091310/WT_/Wellcome Trust/United Kingdom
- RG/09/012/28096/BHF_/British Heart Foundation/United Kingdom
- WT-082597/Z/07/Z/WT_/Wellcome Trust/United Kingdom
- 084183/WT_/Wellcome Trust/United Kingdom
- WT-084183/2/07/2/WT_/Wellcome Trust/United Kingdom
- 082597/WT_/Wellcome Trust/United Kingdom
- RG/09/12/28096/BHF_/British Heart Foundation/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
