

Defects in gallbladder emptying and bile Acid homeostasis in mice with cystic fibrosis transmembrane conductance regulator deficiencies
- PMID: 22370478
- PMCID: PMC3579557
- DOI: 10.1053/j.gastro.2012.02.033
Defects in gallbladder emptying and bile Acid homeostasis in mice with cystic fibrosis transmembrane conductance regulator deficiencies
Abstract
Background & aims: Patients with cystic fibrosis (CF) have poorly defined defects in biliary function. We evaluated the effects of cystic fibrosis transmembrane conductance regulator (CFTR) deficiency on the enterohepatic disposition of bile acids (BAs).
Methods: Bile secretion and BA homeostasis were investigated in Cftr(tm1Unc) (Cftr-/-) and CftrΔF508 (ΔF508) mice.
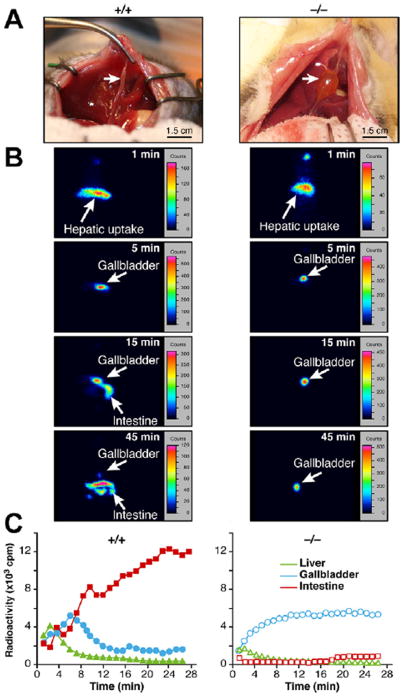
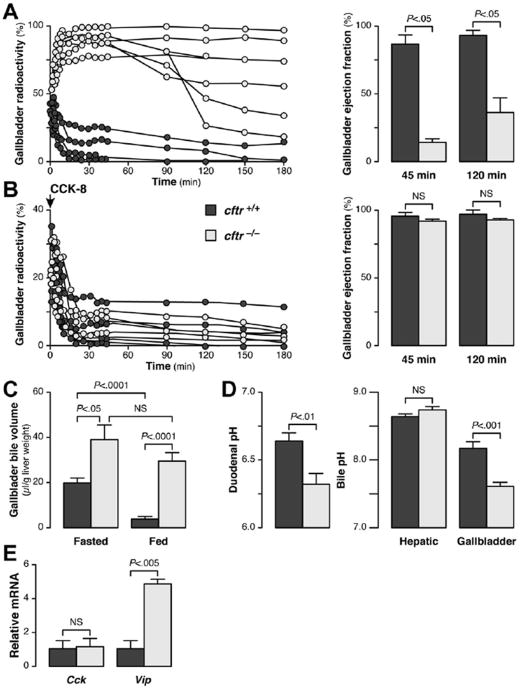
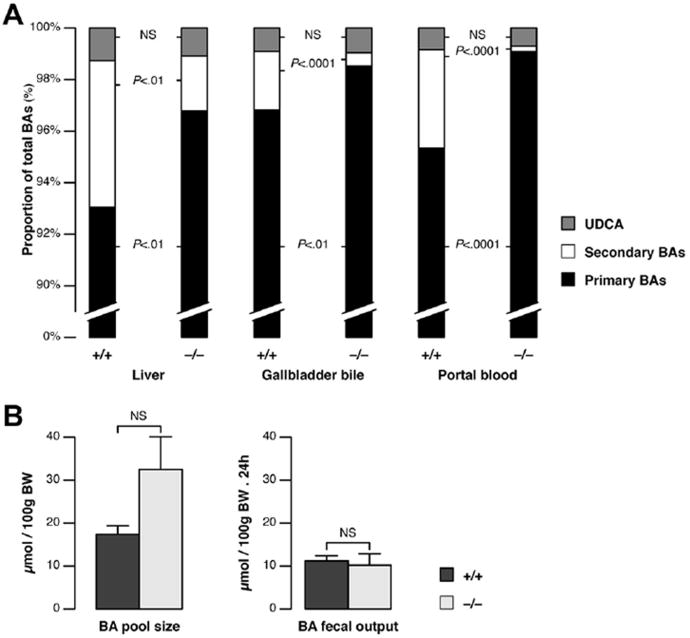
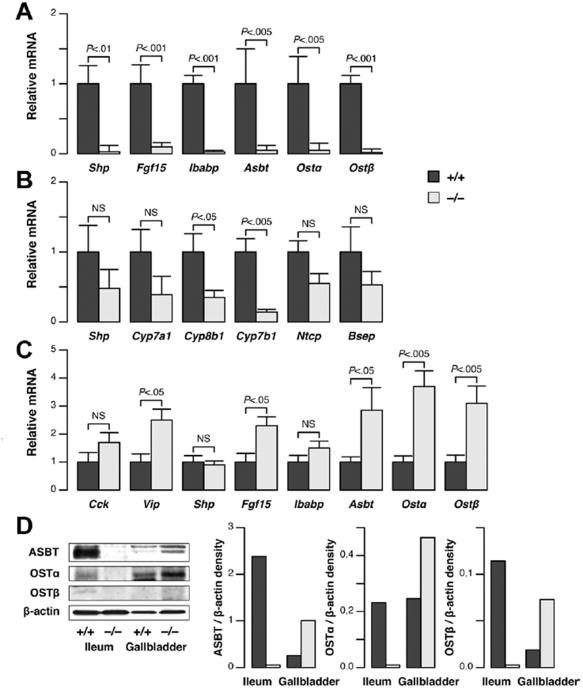
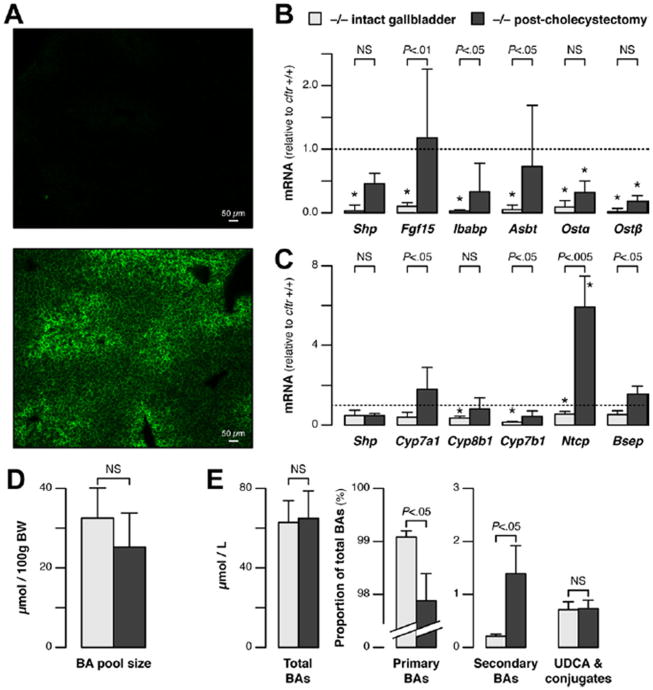
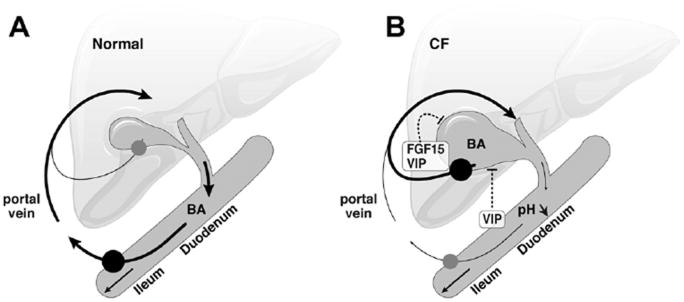
Results: Cftr-/- and ΔF508 mice did not grow to normal size, but did not have liver abnormalities. The gallbladders of Cftr-/- mice were enlarged and had defects in emptying, based on (99m)technetium-mebrofenin scintigraphy or post-prandial variations in gallbladder volume; gallbladder contraction in response to cholecystokinin-8 was normal. Cftr-/- mice had abnormal gallbladder bile and duodenal acidity, and overexpressed the vasoactive intestinal peptide-a myorelaxant factor for the gallbladder. The BA pool was larger in Cftr-/- than wild-type mice, although there were no differences in fecal loss of BAs. Amounts of secondary BAs in portal blood, liver, and bile of Cftr-/- mice were much lower than normal. Expression of genes that are induced by BAs, including fibroblast growth factor-15 and BA transporters, was lower in the ileum but higher in the gallbladders of Cftr-/- mice, compared with wild-type mice, whereas enzymes that synthesize BA were down-regulated in livers of Cftr-/- mice. This indicates that BAs underwent a cholecystohepatic shunt, which was confirmed using cholyl-(Ne-NBD)-lysine as a tracer. In Cftr-/- mice, cholecystectomy reversed most changes in gene expression and partially restored circulating levels of secondary BAs. The ΔF508 mice overexpressed vasoactive intestinal peptide and had defects in gallbladder emptying and in levels of secondary BAs, but these features were less severe than in Cftr-/- mice.
Conclusions: Cftr-/- and CftrΔF508 mice have defects in gallbladder emptying that disrupt enterohepatic circulation of BAs. These defects create a shunt pathway that restricts the amount of toxic secondary BAs that enter the liver.
Copyright © 2012 AGA Institute. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures
Comment in
-
Gallbladder: Cftr affects gallbladder function and bile acid homeostasis.Nat Rev Gastroenterol Hepatol. 2012 Mar 20;9(4):188. doi: 10.1038/nrgastro.2012.46. Nat Rev Gastroenterol Hepatol. 2012. PMID: 22429951 No abstract available.
-
A cholecystohepatic shunt pathway: does the gallbladder protect the liver?Gastroenterology. 2012 Jun;142(7):1416-9. doi: 10.1053/j.gastro.2012.04.036. Epub 2012 Apr 26. Gastroenterology. 2012. PMID: 22542828 No abstract available.
-
Bile acid handling in cystic fibrosis: marked phenotypic differences between mouse models.Gastroenterology. 2012 Dec;143(6):e19-20; author reply e20. doi: 10.1053/j.gastro.2012.08.052. Epub 2012 Oct 19. Gastroenterology. 2012. PMID: 23085355 No abstract available.
References
-
- Hogan DL, Crombie DL, Isenberg JI, et al. Acid-stimulated duodenal bicarbonate secretion involves a CFTR-mediated transport pathway in mice. Gastroenterology. 1997;113:533–541. - PubMed
-
- Chinet T, Fouassier L, Dray-Charier N, et al. Regulation of electrogenic anion secretion in normal and cystic fibrosis gallbladder mucosa. Hepatology. 1999;29:5–13. - PubMed
-
- Zsembery A, Jessner W, Sitter G, et al. Correction of CFTR malfunction and stimulation of Ca-activated Cl channels restore HCO3-secretion in cystic fibrosis bile ductular cells. Hepatology. 2002;35:95–104. - PubMed
-
- Fondacaro JD, Heubi JE, Kellogg FW. Intestinal bile acid malabsorption in cystic fibrosis: a primary mucosal cell defect. Pediatr Res. 1982;16:494–498. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
