

DeSUMOylating isopeptidase: a second class of SUMO protease
- PMID: 22370726
- PMCID: PMC3321169
- DOI: 10.1038/embor.2012.3
DeSUMOylating isopeptidase: a second class of SUMO protease
Abstract
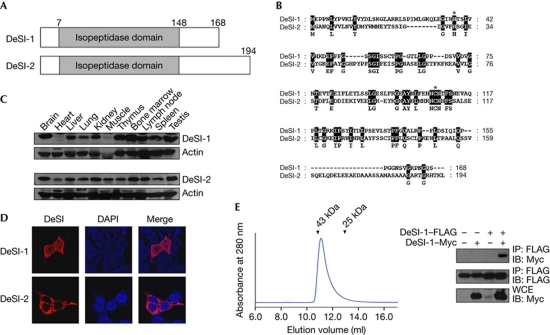
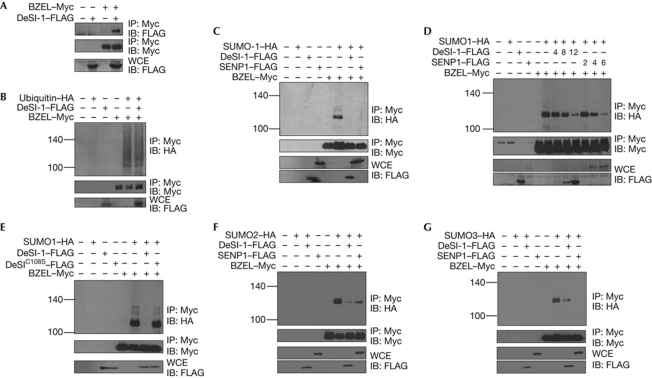
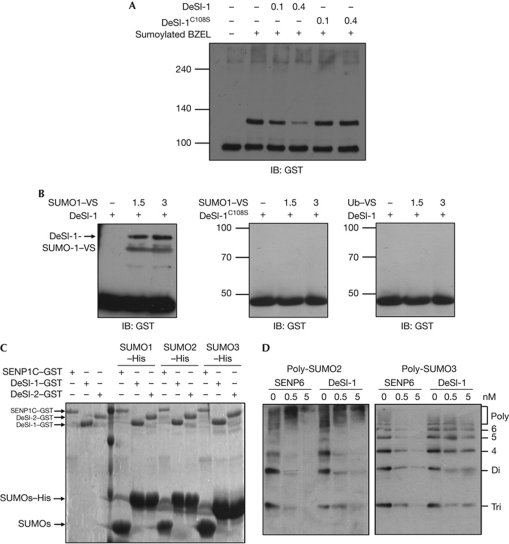
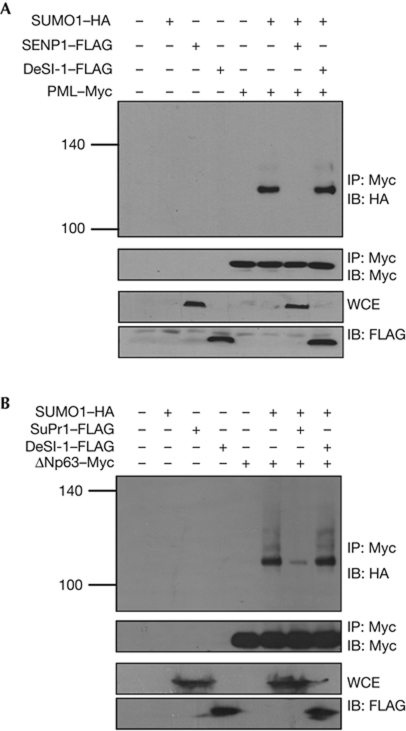
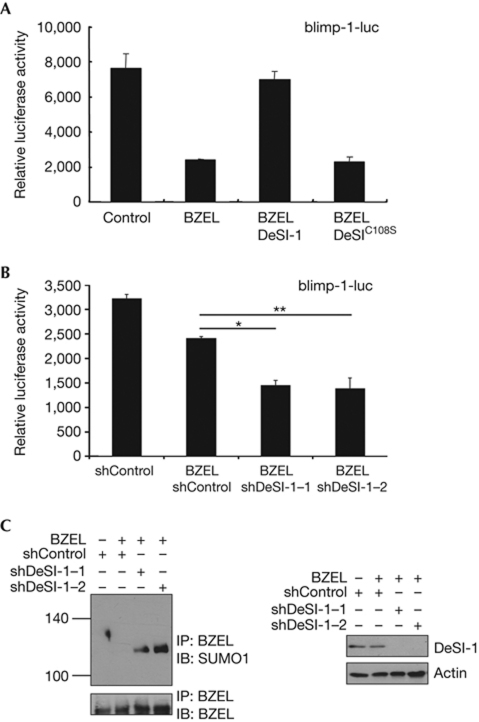
The modification of proteins by small ubiquitin-like modifier (SUMO) is crucial for the regulation of diverse cellular processes. Protein SUMOylation is reversed by isopeptidases, collectively known as deSUMOylases. Only one family of SUMO-specific proteases has been described so far: the sentrin-specific proteases (SENP). Here, we identify and characterize a new deSUMOylase, which we have named DeSI-1 (DeSumoylating Isopeptidase 1). We describe BZEL—a new transcriptional repressor—as substrate of DeSI-1. DeSI-1 catalyses the deSUMOylation, but not the deubiquitination, of BZEL. Furthermore, the SENP substrates PML and ΔNp63 are not deSUMOylated by DeSI-1, suggesting that SENP and DeSI enzymes recognize different sets of substrates. Together, these data identify a second class of SUMO proteases.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Mukhopadhyay D, Dasso M (2007) Modification in reverse: the SUMO proteases. Trends Biochem Sci 32: 286–295 - PubMed
-
- Gill G (2004) SUMO and ubiquitin in the nucleus: different functions, similar mechanisms? Genes Dev 18: 2046–2059 - PubMed
-
- Johnson ES (2004) Protein modification by SUMO. Annu Rev Biochem 73: 355–382 - PubMed
-
- Geiss-Friedlander R, Melchior F (2007) Concepts in sumoylation: a decade on. Nat Rev Mol Cell Biol 8: 947–956 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
