

Consensus paper: pathological role of the cerebellum in autism
- PMID: 22370873
- PMCID: PMC3677555
- DOI: 10.1007/s12311-012-0355-9
Consensus paper: pathological role of the cerebellum in autism
Abstract
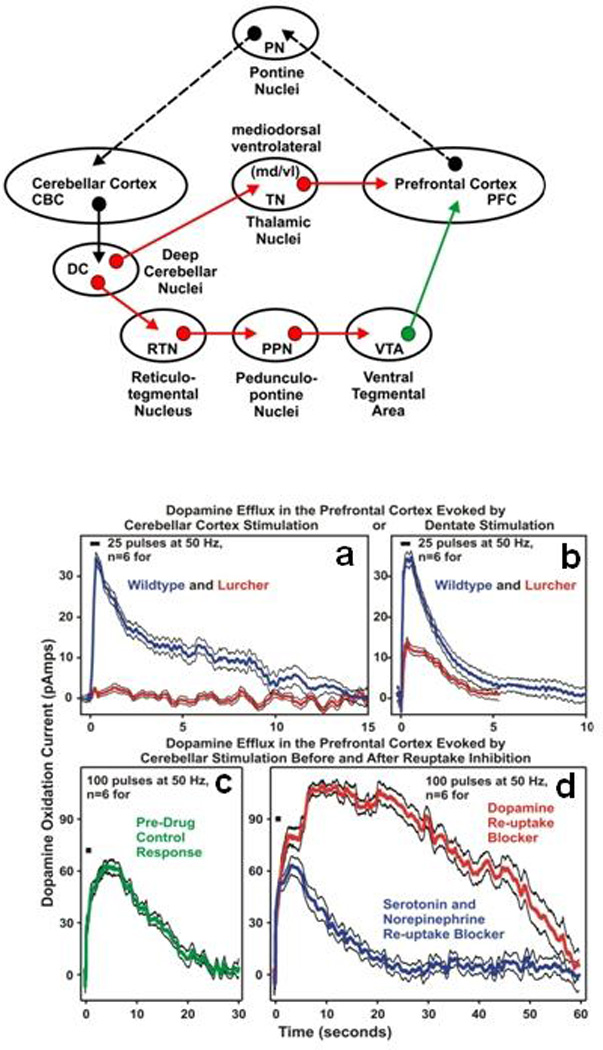
There has been significant advancement in various aspects of scientific knowledge concerning the role of cerebellum in the etiopathogenesis of autism. In the current consensus paper, we will observe the diversity of opinions regarding the involvement of this important site in the pathology of autism. Recent emergent findings in literature related to cerebellar involvement in autism are discussed, including: cerebellar pathology, cerebellar imaging and symptom expression in autism, cerebellar genetics, cerebellar immune function, oxidative stress and mitochondrial dysfunction, GABAergic and glutamatergic systems, cholinergic, dopaminergic, serotonergic, and oxytocin-related changes in autism, motor control and cognitive deficits, cerebellar coordination of movements and cognition, gene-environment interactions, therapeutics in autism, and relevant animal models of autism. Points of consensus include presence of abnormal cerebellar anatomy, abnormal neurotransmitter systems, oxidative stress, cerebellar motor and cognitive deficits, and neuroinflammation in subjects with autism. Undefined areas or areas requiring further investigation include lack of treatment options for core symptoms of autism, vermal hypoplasia, and other vermal abnormalities as a consistent feature of autism, mechanisms underlying cerebellar contributions to cognition, and unknown mechanisms underlying neuroinflammation.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures
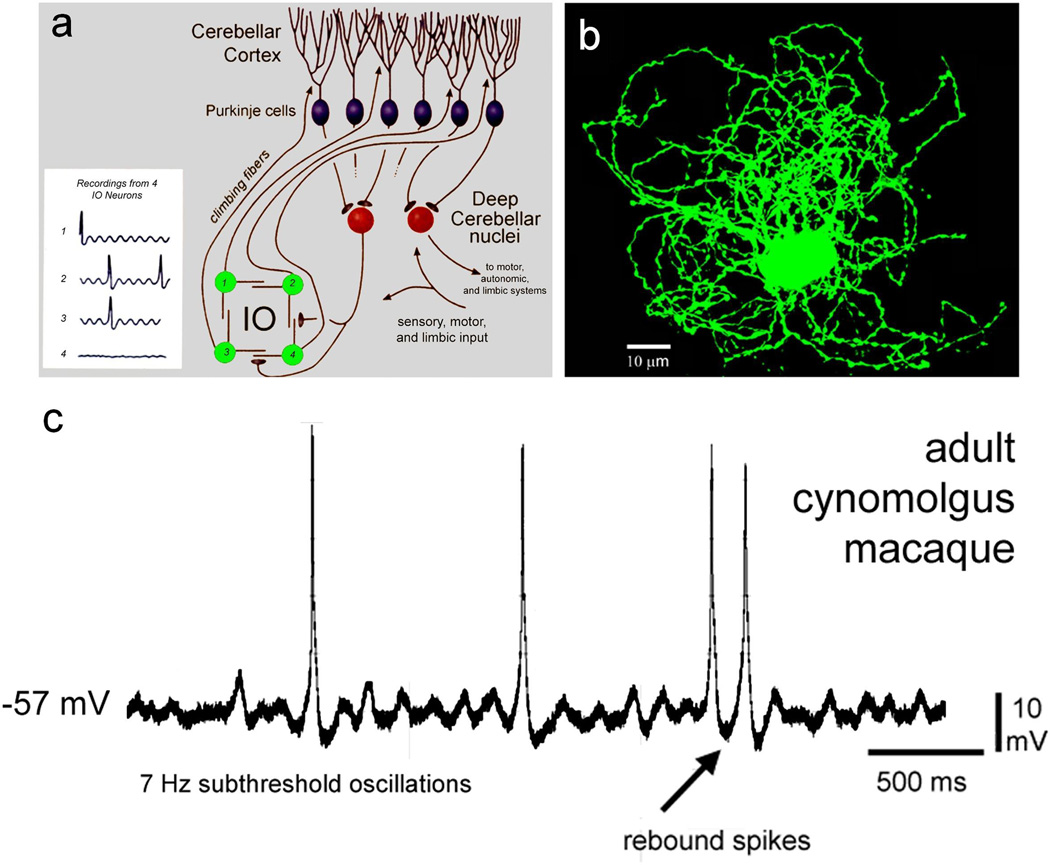
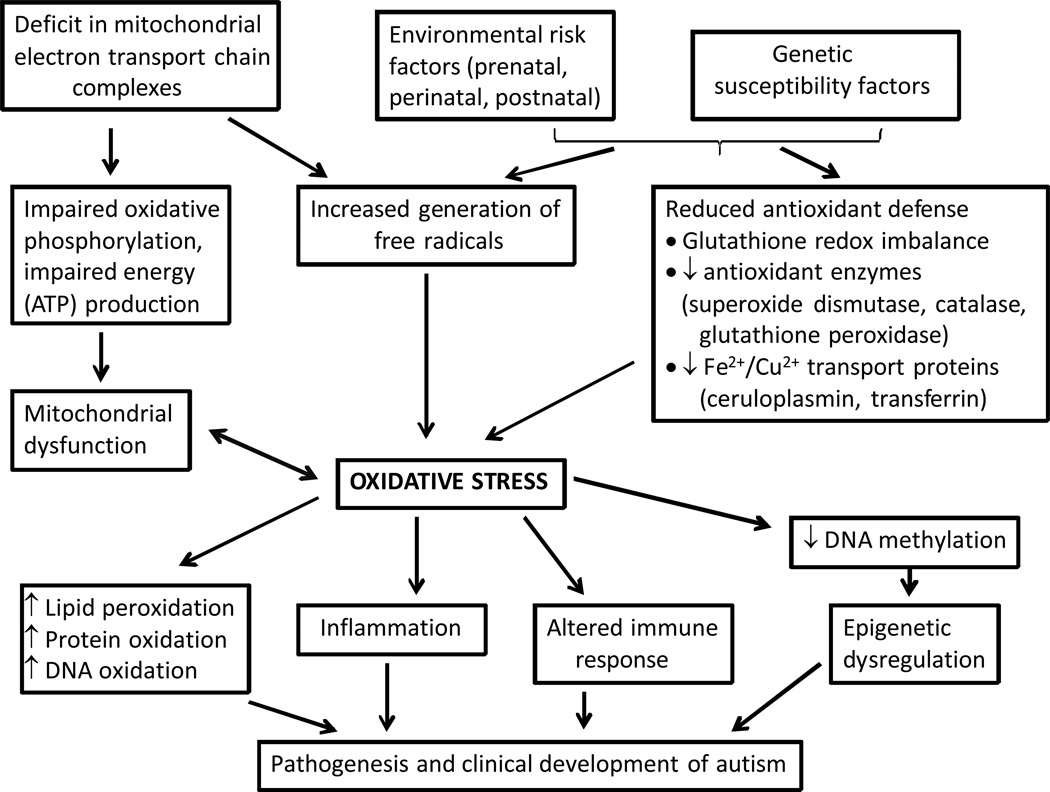
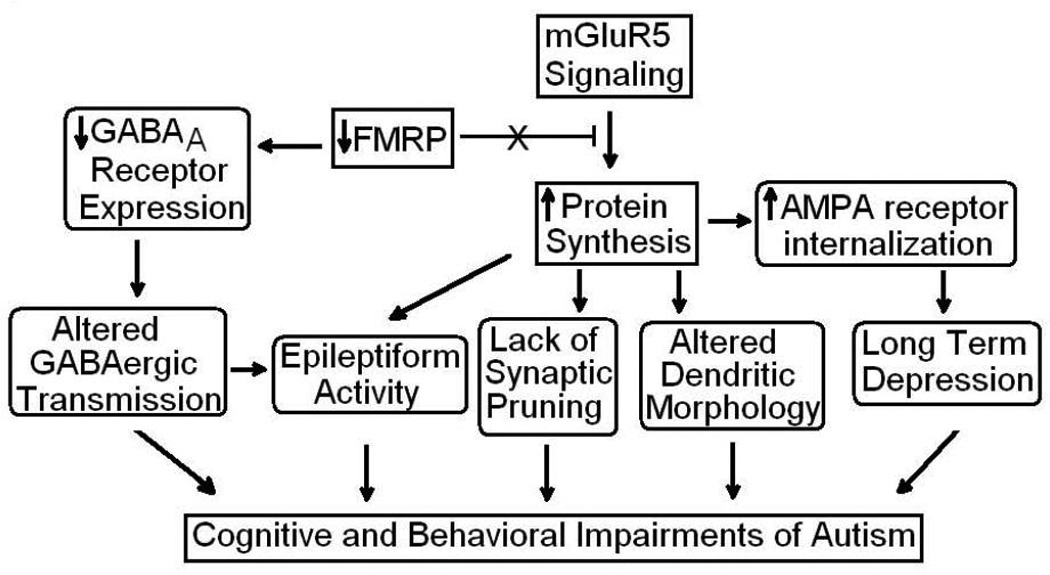
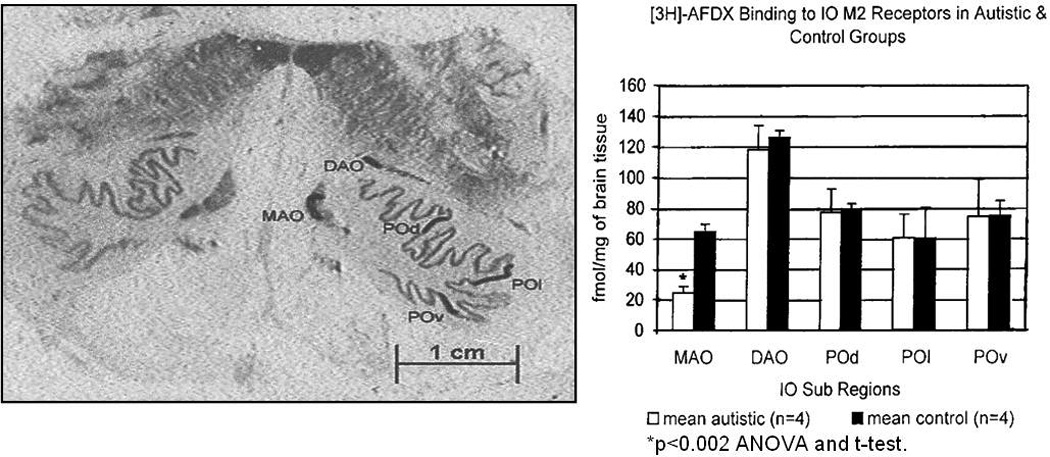
Comment in
-
The cerebellum in autism: pathogenic or an anatomical beacon?Cerebellum. 2013 Oct;12(5):776-7. doi: 10.1007/s12311-013-0483-x. Cerebellum. 2013. PMID: 23605119 Free PMC article. No abstract available.
-
Cerebellum and autism.Cerebellum. 2013 Oct;12(5):778-9. doi: 10.1007/s12311-013-0484-9. Cerebellum. 2013. PMID: 23605188 No abstract available.
References
-
- American Psychiatric Association Diagnostic and Statistical Manuel of Mental Disorders. (DSM-4) 4th ed. Washington, DC: APA; 1994.
-
- Wassink TH, Brzustowicz LM, Bartlett CW, Szatmari P. The search for autism disease genes. Ment Retard Dev Disabil Res Rev. 2004;10:272–283. - PubMed
-
- Bauman ML, Kemper TL. Histoanatomic observations of the brain in early infantile autism. Neurology. 1985;35:866–874. - PubMed
-
- Arin DM, Bauman ML, Kemper TL. The distribution of Purkinje cell loss in the cerebellum in autism. Neurology. 1991;41(Suppl):307.
-
- Bailey A, Luthert P, Dean A, Harding B, Janota I, Montgomery M, et al. A clinicopathological study of autism. Brain. 1998;121:889–905. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P50 HD055782/HD/NICHD NIH HHS/United States
- R01-HD055741/HD/NICHD NIH HHS/United States
- P50HD055751/HD/NICHD NIH HHS/United States
- 3R01HD05207403-S1/HD/NICHD NIH HHS/United States
- K23 MH092696/MH/NIMH NIH HHS/United States
- R01NS060887/NS/NINDS NIH HHS/United States
- R01NS063009/NS/NINDS NIH HHS/United States
- R01-NS31224-18/NS/NINDS NIH HHS/United States
- 5R01HD052074-05/HD/NICHD NIH HHS/United States
- HD35465/HD/NICHD NIH HHS/United States
- R01 NS063009/NS/NINDS NIH HHS/United States
- KL2 TR000131/TR/NCATS NIH HHS/United States
- P01 HD035465/HD/NICHD NIH HHS/United States
- AS3587/AS/Autism Speaks/United States
- R01-HD065283/HD/NICHD NIH HHS/United States
- R01 NS060887/NS/NINDS NIH HHS/United States
- R21HD065269/HD/NICHD NIH HHS/United States
- P50-HD055782/HD/NICHD NIH HHS/United States
- R01 HD052074/HD/NICHD NIH HHS/United States
- R21 HD065269/HD/NICHD NIH HHS/United States
- R01 NS038975/NS/NINDS NIH HHS/United States
- P50 HD055751/HD/NICHD NIH HHS/United States
- NS38975-05/NS/NINDS NIH HHS/United States
- 1K23MH092696/MH/NIMH NIH HHS/United States
- R01 NS031224/NS/NINDS NIH HHS/United States
- R01 HD05571/HD/NICHD NIH HHS/United States
- R01 HD039459/HD/NICHD NIH HHS/United States
- R01 NS31224-18/NS/NINDS NIH HHS/United States
- R01 NS072441/NS/NINDS NIH HHS/United States
- R01 HD065283/HD/NICHD NIH HHS/United States
- UL1 TR000130/TR/NCATS NIH HHS/United States
- 5R01HD039459-06/HD/NICHD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
