

Estradiol increases hematopoietic stem and progenitor cells independent of its actions on bone
- PMID: 22371175
- PMCID: PMC3409808
- DOI: 10.3324/haematol.2011.052456
Estradiol increases hematopoietic stem and progenitor cells independent of its actions on bone
Abstract
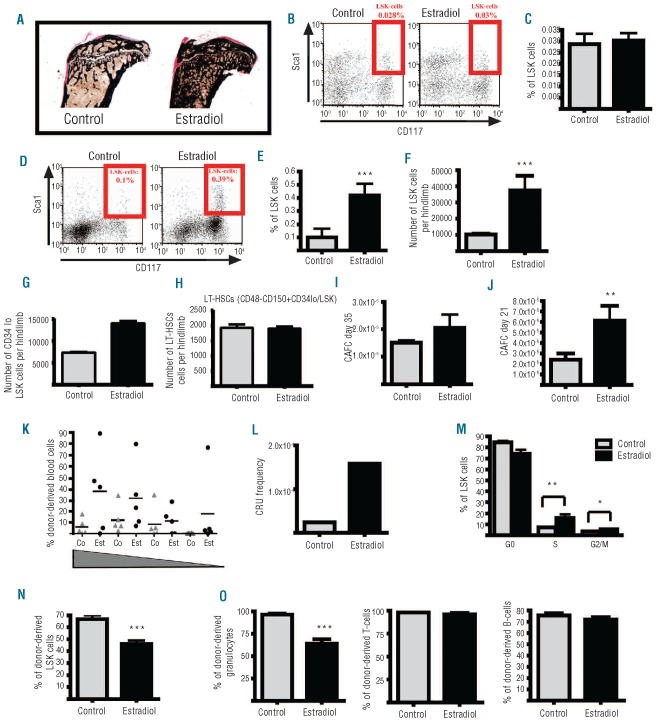
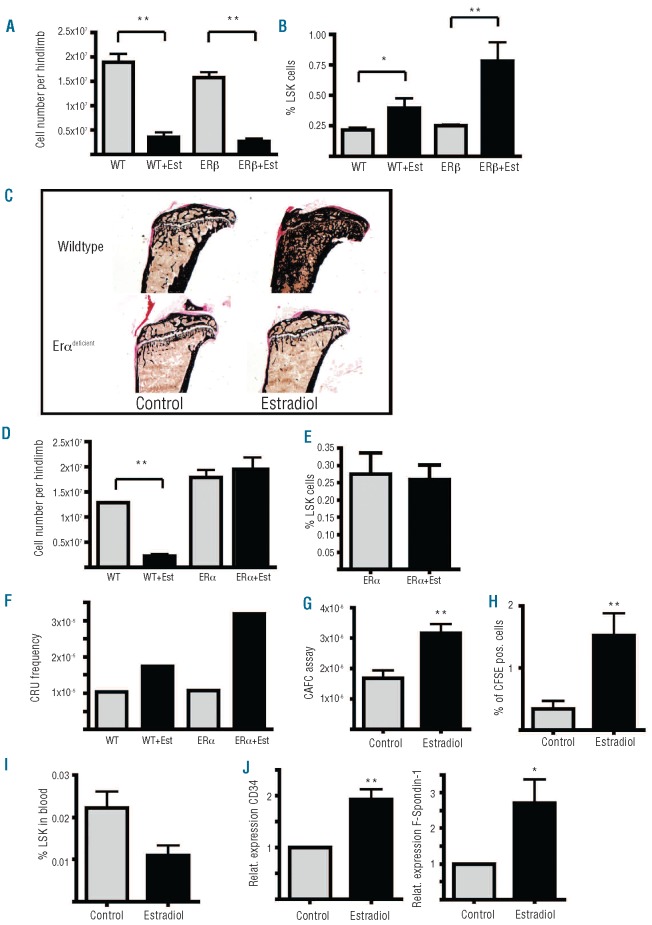
Hematopoietic stem and progenitor cells reside in vascular and endosteal niches in the bone marrow. Factors affecting bone remodeling were reported to influence numbers and mobilization of hematopoietic stem cells. We therefore analyzed the effects of estradiol acting anabolic on bone integrity. Here we observe that estradiol increases progenitor cell numbers in the vascular but not in the endosteal compartment independent of its estrogen receptor α-dependent anabolic bone effects. Hematopoietic progenitors capable of reconstituting lethally irradiated mice are increased by enhanced cell cycle entry, leading to a diminished long-term reconstitution potential after serial transplantation. We demonstrate that estradiol action on stromal cells potently favors hematopoietic progenitor/stem cell frequency accompanied by enhanced expression of cell adhesion molecules. Finally, estradiol treatment enhances retention of hematopoietic stem cells in the vascular niche of the bone marrow. We describe for the first time the mechanism of estrogen action on hematopoietic stem and progenitor cells.
Figures
References
-
- Fuchs E, Tumbar T, Guasch G. Socializing with the neighbors: stem cells and their niche. Cell. 2004;116(6):769–78. - PubMed
-
- Sugiyama T, Kohara H, Noda M, Nagasawa T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity. 2006;25(6):977–88. - PubMed
-
- Calvi LM, Adams GB, Weibrecht KW, Weber JM, Olson DP, Knight MC, et al. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature. 2003;425(6960):841–6. - PubMed
-
- Zhang J, Niu C, Ye L, Huang H, He X, Tong WG, et al. Identification of the haematopoietic stem cell niche and control of the niche size. Nature. 2003;425(6960):836–41. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
