

Structural basis for dual nucleotide selectivity of aminoglycoside 2''-phosphotransferase IVa provides insight on determinants of nucleotide specificity of aminoglycoside kinases
- PMID: 22371504
- PMCID: PMC3339986
- DOI: 10.1074/jbc.M112.349670
Structural basis for dual nucleotide selectivity of aminoglycoside 2''-phosphotransferase IVa provides insight on determinants of nucleotide specificity of aminoglycoside kinases
Abstract
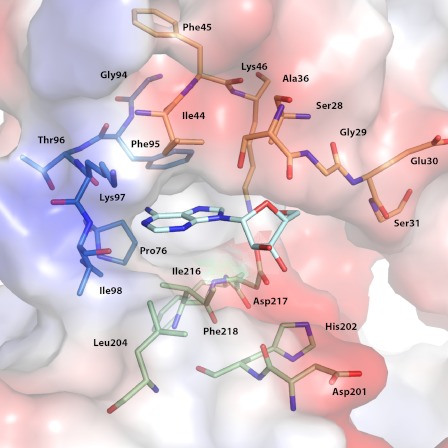
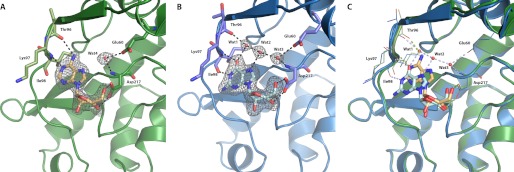
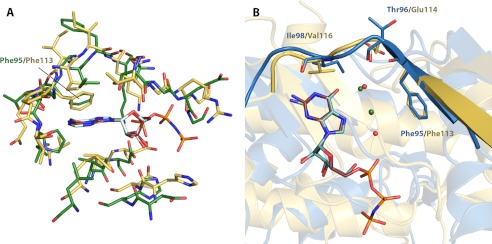
Enzymatic phosphorylation through a family of enzymes called aminoglycoside O-phosphotransferases (APHs) is a major mechanism by which bacteria confer resistance to aminoglycoside antibiotics. Members of the APH(2″) subfamily are of particular clinical interest because of their prevalence in pathogenic strains and their broad substrate spectra. APH(2″) enzymes display differential preferences between ATP or GTP as the phosphate donor, with aminoglycoside 2″-phosphotransferase IVa (APH(2″)-IVa) being a member that utilizes both nucleotides at comparable efficiencies. We report here four crystal structures of APH(2″)-IVa, two of the wild type enzyme and two of single amino acid mutants, each in complex with either adenosine or guanosine. Together, these structures afford a detailed look at the nucleoside-binding site architecture for this enzyme and reveal key elements that confer dual nucleotide specificity, including a solvent network in the interior of the nucleoside-binding pocket and the conformation of an interdomain linker loop. Steady state kinetic studies, as well as sequence and structural comparisons with members of the APH(2″) subfamily and other aminoglycoside kinases, rationalize the different substrate preferences for these enzymes. Finally, despite poor overall sequence similarity and structural homology, analysis of the nucleoside-binding pocket of APH(2″)-IVa shows a striking resemblance to that of eukaryotic casein kinase 2 (CK2), which also exhibits dual nucleotide specificity. These results, in complement with the multitude of existing inhibitors against CK2, can serve as a structural basis for the design of nucleotide-competitive inhibitors against clinically relevant APH enzymes.
Figures


Similar articles
-
Revisiting the nucleotide and aminoglycoside substrate specificity of the bifunctional aminoglycoside acetyltransferase(6')-Ie/aminoglycoside phosphotransferase(2'')-Ia enzyme.J Biol Chem. 2012 Dec 21;287(52):43262-9. doi: 10.1074/jbc.M112.416453. Epub 2012 Oct 31. J Biol Chem. 2012. PMID: 23115238 Free PMC article.
-
Aminoglycoside 2''-phosphotransferase IIIa (APH(2'')-IIIa) prefers GTP over ATP: structural templates for nucleotide recognition in the bacterial aminoglycoside-2'' kinases.J Biol Chem. 2012 Apr 13;287(16):12893-903. doi: 10.1074/jbc.M112.341206. Epub 2012 Feb 24. J Biol Chem. 2012. PMID: 22367198 Free PMC article.
-
Crystal structure and kinetic mechanism of aminoglycoside phosphotransferase-2''-IVa.Protein Sci. 2010 Aug;19(8):1565-76. doi: 10.1002/pro.437. Protein Sci. 2010. PMID: 20556826 Free PMC article.
-
Developing a snapshot of the ATP binding domain(s) of aminoglycoside phosphotransferases.Front Biosci. 1999 Jan 1;4:D63-71. doi: 10.2741/perlin. Front Biosci. 1999. PMID: 9872732 Review.
-
Aminoglycoside phosphotransferases: proteins, structure, and mechanism.Front Biosci. 1999 Jan 1;4:D9-21. doi: 10.2741/wright. Front Biosci. 1999. PMID: 9872733 Review.
Cited by
-
Prospects for circumventing aminoglycoside kinase mediated antibiotic resistance.Front Cell Infect Microbiol. 2013 Jun 25;3:22. doi: 10.3389/fcimb.2013.00022. eCollection 2013. Front Cell Infect Microbiol. 2013. PMID: 23805415 Free PMC article. Review.
-
Discovery and characterization of genes conferring natural resistance to the antituberculosis antibiotic capreomycin.Commun Biol. 2023 Dec 19;6(1):1282. doi: 10.1038/s42003-023-05681-6. Commun Biol. 2023. PMID: 38114770 Free PMC article.
-
APH Inhibitors that Reverse Aminoglycoside Resistance in Enterococcus casseliflavus.ChemMedChem. 2025 Apr 14;20(8):e202400842. doi: 10.1002/cmdc.202400842. Epub 2025 Jan 26. ChemMedChem. 2025. PMID: 39801466 Free PMC article.
-
Bulky "gatekeeper" residue changes the cosubstrate specificity of aminoglycoside 2''-phosphotransferase IIa.Antimicrob Agents Chemother. 2013 Aug;57(8):3763-6. doi: 10.1128/AAC.00381-13. Epub 2013 May 28. Antimicrob Agents Chemother. 2013. PMID: 23716051 Free PMC article.
-
Antibiotic resistance and inhibition mechanism of novel aminoglycoside phosphotransferase APH(5) from B. subtilis subsp. subtilis strain RK.Braz J Microbiol. 2019 Oct;50(4):887-898. doi: 10.1007/s42770-019-00132-z. Epub 2019 Aug 10. Braz J Microbiol. 2019. PMID: 31401782 Free PMC article.
References
-
- Smith C. A., Baker E. N. (2002) Aminoglycoside antibiotic resistance by enzymatic deactivation. Curr. Drug Targets Infect. Disord. 2, 143–160 - PubMed
-
- Chow J. W. (2000) Aminoglycoside resistance in enterococci. Clin. Infect. Dis. 31, 586–589 - PubMed
-
- Burk D. L., Berghuis A. M. (2002) Protein kinase inhibitors and antibiotic resistance. Pharmacol. Ther. 93, 283–292 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
