

RNase H-mediated degradation of toxic RNA in myotonic dystrophy type 1
- PMID: 22371589
- PMCID: PMC3306674
- DOI: 10.1073/pnas.1117019109
RNase H-mediated degradation of toxic RNA in myotonic dystrophy type 1
Abstract
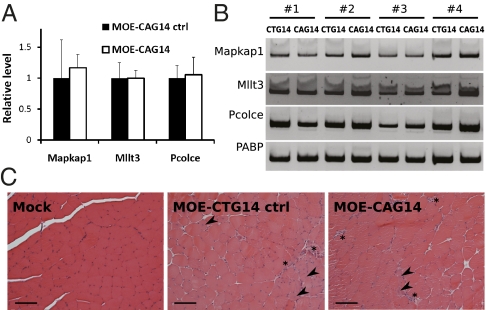
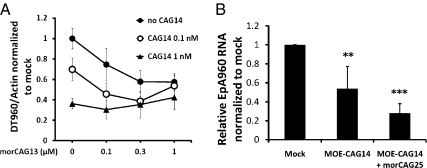
Myotonic dystrophy type 1 (DM1) is an RNA-dominant disease caused by abnormal transcripts containing expanded CUG repeats. The CUG transcripts aggregate in the nucleus to form RNA foci and lead to nuclear depletion of Muscleblind-like 1 (MBNL1) and stabilized expression of CUGBP Elav like family 1 (CELF1), both of which are splicing regulatory proteins. The imbalance of these proteins results in misregulation of alternative splicing and neuromuscular abnormalities. Here, we report the use of antisense oligonucleotides (ASOs) as a therapeutic approach to target the pathogenic RNA in DM1. We designed chimeric ASOs, termed gapmers, containing modified nucleic acid residues to induce RNase H-mediated degradation of CUG-repeat transcripts. The gapmers selectively knockdown expanded CUG transcripts and are sufficient to disrupt RNA foci both in cell culture and mouse models for DM1. Furthermore, combination of gapmers with morpholino ASOs that help release binding of MBNL1 to the toxic RNA can potentially enhance the knockdown effect. Additional optimization will be required for systemic delivery; however, our study provides an alternative strategy for the use of ASOs in DM1 therapy.
Conflict of interest statement
Conflict of interest statement: C.F.B. is an employee of Isis Pharmaceuticals, Inc., and may materially benefit either directly or indirectly through stock options.
Figures
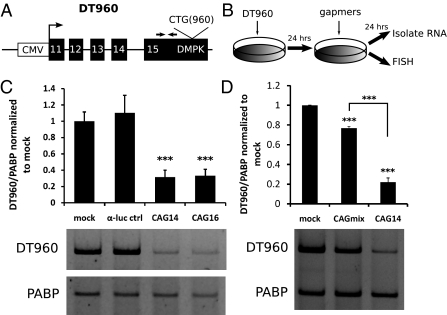
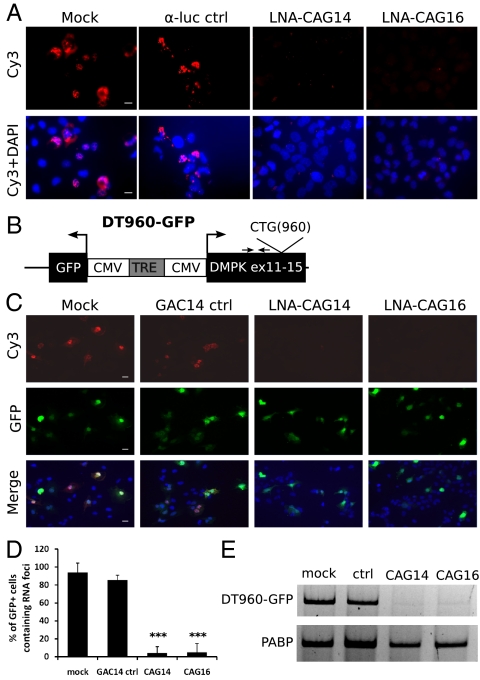
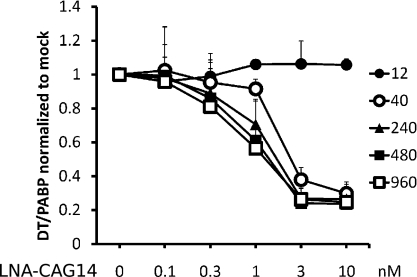
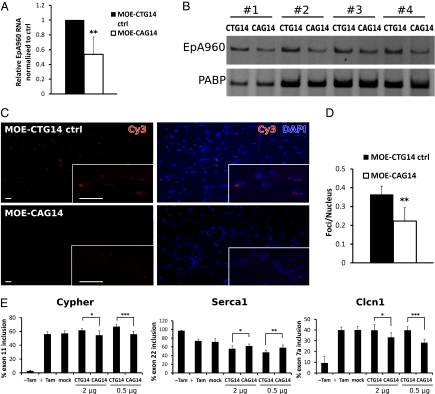
References
-
- Harper P.S. Myotonic Dystrophy. London: W.B. Saunders; 2001.
-
- Brook JD, et al. Molecular basis of myotonic dystrophy: expansion of a trinucleotide (CTG) repeat at the 3′ end of a transcript encoding a protein kinase family member. Cell. 1992;68:799–808. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
