

Transcriptome sequencing identifies SPL7-regulated copper acquisition genes FRO4/FRO5 and the copper dependence of iron homeostasis in Arabidopsis
- PMID: 22374396
- PMCID: PMC3315244
- DOI: 10.1105/tpc.111.090431
Transcriptome sequencing identifies SPL7-regulated copper acquisition genes FRO4/FRO5 and the copper dependence of iron homeostasis in Arabidopsis
Abstract
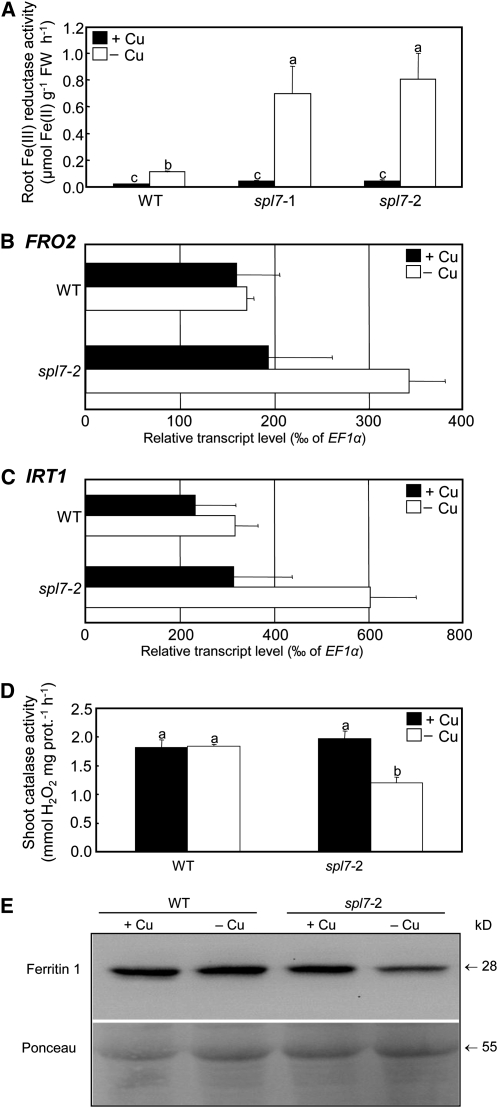
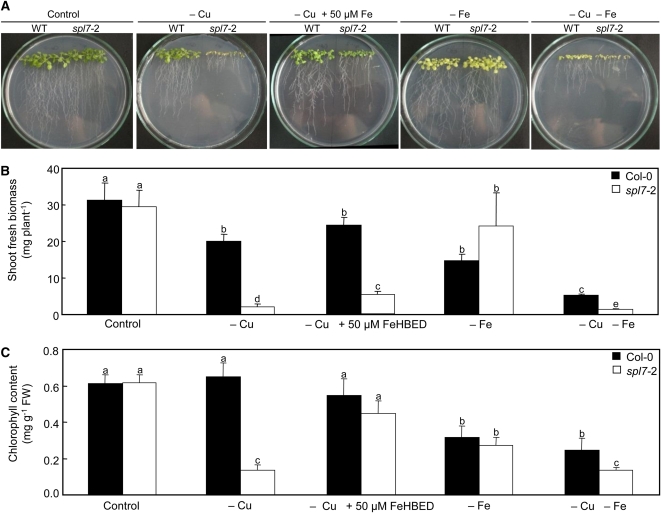
The transition metal copper (Cu) is essential for all living organisms but is toxic when present in excess. To identify Cu deficiency responses comprehensively, we conducted genome-wide sequencing-based transcript profiling of Arabidopsis thaliana wild-type plants and of a mutant defective in the gene encoding SQUAMOSA PROMOTER BINDING PROTEIN-LIKE7 (SPL7), which acts as a transcriptional regulator of Cu deficiency responses. In response to Cu deficiency, FERRIC REDUCTASE OXIDASE5 (FRO5) and FRO4 transcript levels increased strongly, in an SPL7-dependent manner. Biochemical assays and confocal imaging of a Cu-specific fluorophore showed that high-affinity root Cu uptake requires prior FRO5/FRO4-dependent Cu(II)-specific reduction to Cu(I) and SPL7 function. Plant iron (Fe) deficiency markers were activated in Cu-deficient media, in which reduced growth of the spl7 mutant was partially rescued by Fe supplementation. Cultivation in Cu-deficient media caused a defect in root-to-shoot Fe translocation, which was exacerbated in spl7 and associated with a lack of ferroxidase activity. This is consistent with a possible role for a multicopper oxidase in Arabidopsis Fe homeostasis, as previously described in yeast, humans, and green algae. These insights into root Cu uptake and the interaction between Cu and Fe homeostasis will advance plant nutrition, crop breeding, and biogeochemical research.
Figures







References
-
- Abdel-Ghany S.E. (2009). Contribution of plastocyanin isoforms to photosynthesis and copper homeostasis in Arabidopsis thaliana grown at different copper regimes. Planta 229: 767–779 - PubMed
-
- Allison D.B., Gadbury G.L., Heo M.S., Fernández J.R., Lee C.K., Prolla T.A., Weindruch R. (2002). A mixture model approach for the analysis of microarray gene expression data. Comput. Stat. Data Anal. 39: 1–20
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
