

Epigenetic disruption of cadherin-11 in human cancer metastasis
- PMID: 22374749
- PMCID: PMC3467766
- DOI: 10.1002/path.4011
Epigenetic disruption of cadherin-11 in human cancer metastasis
Abstract
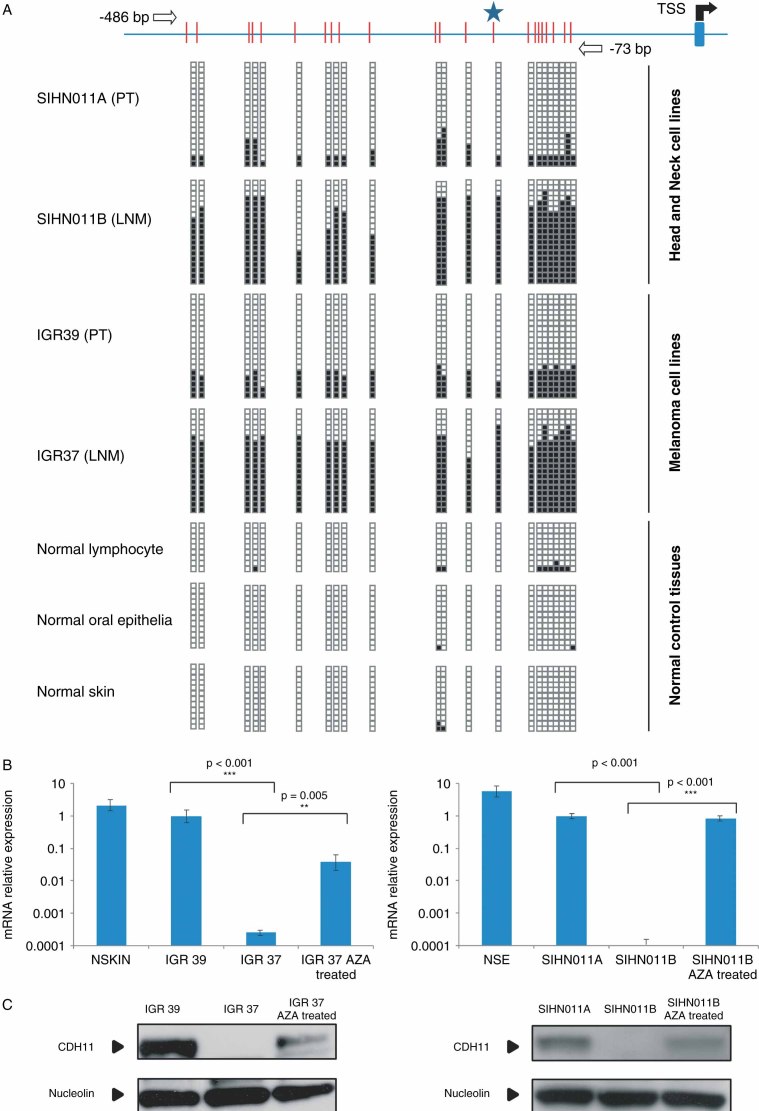
Little is known about the molecular events occurring in the metastases of human tumours. Epigenetic alterations are dynamic lesions that change over the natural course of the disease, and so they might play a role in the biology of cancer cells that have departed from the primary tumour. Herein, we have adopted an epigenomic approach to identify some of these changes. Using a DNA methylation microarray platform to compare paired primary tumour and lymph node metastatic cell lines from the same patient, we observed cadherin-11 promoter CpG island hypermethylation as a likely target of the process. We found that CDH11 DNA methylation-associated transcriptional silencing occurred in the corresponding lymph node metastases of melanoma and head and neck cancer cells but not in the primary tumours. Using in vitro and in vivo cellular and mouse models for depleted or enhanced CDH11 activity, we also demonstrated that CDH11 acts as an inhibitor of tumour growth, motility and dissemination. Most importantly, the study of CDH11 5'-CpG island hypermethylation in primary tumours and lymph node metastases of cancer patients showed this epigenetic alteration to be significantly confined to the disseminated cells. Overall, these results indicate the existence of metastasis-specific epigenetic events that might contribute to the progression of the disease.
Copyright © 2012 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
Figures




References
-
- Jemal A, Siegel R, Xu J, et al. Cancer statistics, 2010. CA Cancer J Clin. 2010;60:277–300. - PubMed
-
- Fidler IJ. The pathogenesis of cancer metastasis: The ‘seed and soil’ hypothesis revisited. Nat Rev Cancer. 2003;3:453–458. - PubMed
-
- Carmona FJ, Esteller M. IDIBELL cancer conference on metastasis and angiogenesis. Cancer Res. 2011;71:6097–7101. - PubMed
-
- Ma L, Teruya-Feldstein J, Weinberg RA. Tumour invasion and metastasis initiated by microRNA-10b in breast cancer. Nature. 2007;449:682–688. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
