

Comment
. 2012 Apr;135(Pt 4):e215; author reply e216.
doi: 10.1093/brain/aws017.
Epub 2012 Feb 28.
Harnessing neuroplasticity for clinical applications
- PMID: 22374936
- PMCID: PMC3326250
- DOI: 10.1093/brain/aws017
Item in Clipboard
Comment
Harnessing neuroplasticity for clinical applications
Brain.
2012 Apr.
No abstract available
Figures
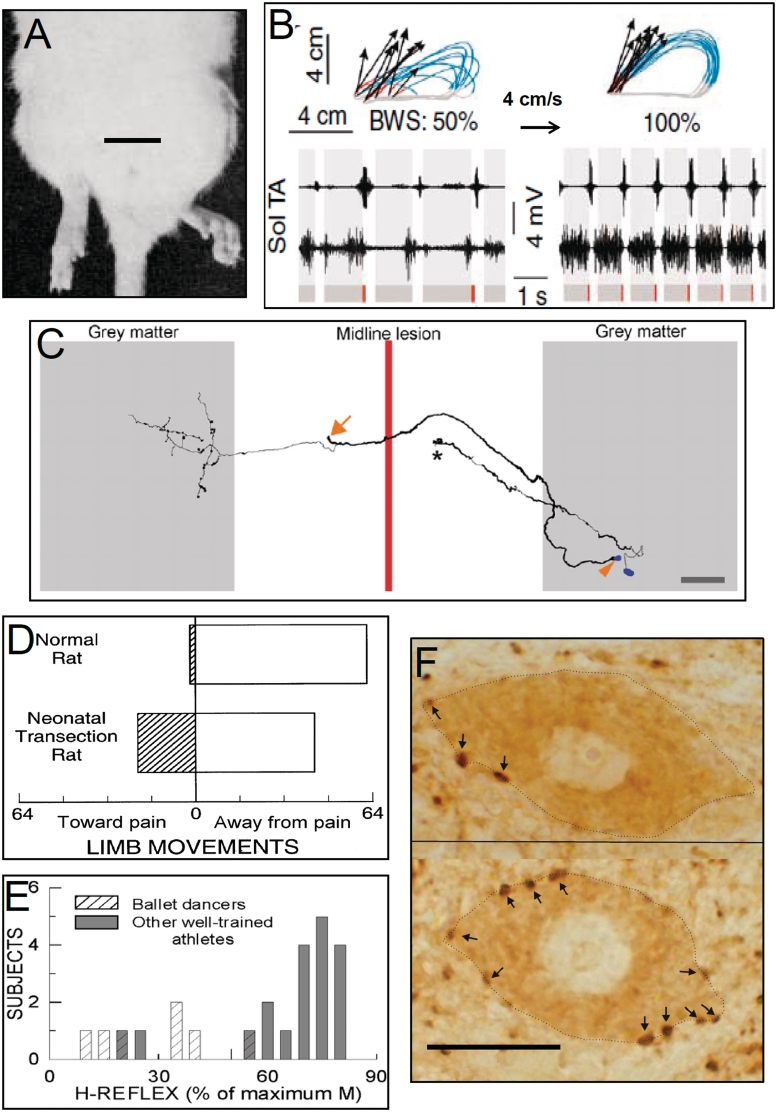
Six representative examples of activity-dependent spinal cord plasticity. The first three (A–C) illustrate spinal cord plasticity in pathological situations or in reduced preparations. The next three (D–F) illustrate spinal cord plasticity in normal life. (A) A hindlimb postural asymmetry produced by a unilateral cerebellar lesion persists after complete transection of the thoracic spinal cord. The cerebellar lesion occurred 60 min prior to the spinal cord transection. Scale bar = 2 cm. Modified from Chamberlain et al. (1963). (B) Impact of a combined treatment of serotonergic agonists, epidural electrical stimulation and locomotor training on treadmill locomotion (i.e. at 4 cm/s) in spinal-transected rats. Data from an untreated and a treated rat are shown on the left and right, respectively. Top: a stick diagram decomposition of hindlimb motion during swing. Also shown are limb end-point trajectories (with red indicating the initial drag phase of swing) and vectors representing the direction and magnitude of limb end-point velocity at swing onset. The rat's per cent of body weight support (BWS) is indicated. Bottom: sequences of EMG activity from tibialis anterior (TA) and soleus (Sol) muscles. Grey and red bars indicate the stance and drag phases, respectively. Locomotion is far more normal, effective and consistent in the treated rat. Modified from Courtine et al. (2009). (C) Reconstruction of a propriospinal commissural interneuron in the rat spinal cord that has regenerated through a midline lesion made 72 days earlier and formed collaterals. The arrowhead indicates the soma of this neuron. The regenerated axons of such neurons make functional synapses. The arrow marks the place where the regenerating axon bifurcated and continued rostrally and caudally (branches not shown for clarity). Also shown is another neuron that has not regenerated across the lesion and terminates in a growth cone (asterisk). Scale bar = 200 μm. From Fenrich and Rose (2009). (D) The direction of flexion withdrawal responses to painful stimuli in normal adult rats and in adult rats in which the spinal cord was transected just after birth. In normal adults, the direction of the response is almost always correct (i.e. the limb moves away from the painful stimulus), while in transected adults it is often incorrect (i.e. the limb moves towards the stimulus). Neonatal spinal cord transection abolishes the descending input that gradually shapes normal (i.e. correct) flexion withdrawal responses. Modified from Levinsson et al. (1999). (E) Soleus H-reflexes are much smaller in professional ballet dancers than in other well-trained athletes (e.g. runners, swimmers and cyclists). (H-reflexes of sedentary subjects fall in between.) The dancers' smaller reflexes appear to be an important component of skill acquisition. Modified from Nielsen et al. (1993). (F) Soleus motor neurons (dotted lines) from a control rat (top) and a rat in which the soleus H-reflex was reduced by an operant down-conditioning protocol (bottom). Arrows point to GABAergic terminals on the somatic membrane. The terminals are identified by glutamic acid decarboxylase (GAD67)-immunoreactivity. After down-conditioning, soleus motor neurons have many more GABAergic terminals, and these terminals are more densely labelled and cover more of the somatic membrane. The increase in GABAergic terminals is likely to be a component of the spinal cord plasticity that produces the smaller H-reflex. Scale bar = 20 μm. See Wang et al. (2006) for full information.
Comment on
-
Harnessing neuroplasticity for clinical applications.Brain. 2011 Jun;134(Pt 6):1591-609. doi: 10.1093/brain/awr039. Epub 2011 Apr 10. Brain. 2011. PMID: 21482550 Free PMC article.
References
-
- Brodal A. Neurological anatomy in relation to clinical medicine. New York/Oxford: Oxford University Press; 1981.
-
- Carp JS, Wolpaw JR. Motoneuron plasticity underlying operantly conditioned decrease in primate H-reflex. J Neurophysiol. 1994;72:431–42. - PubMed
-
- Carp JS, Chen XY, Sheikh H, Wolpaw JR. Operant conditioning of rat H-reflex affects motoneuron axonal conduction velocity. Exp Br Res. 2001;136:269–73. - PubMed
-
- Chamberlain T, Halick P, Gerard RW. Fixation of experience in the rat spinal cord. J. Neurophysiol. 1963;22:662–73. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
