

Cellulose microfibril crystallinity is reduced by mutating C-terminal transmembrane region residues CESA1A903V and CESA3T942I of cellulose synthase
- PMID: 22375033
- PMCID: PMC3306678
- DOI: 10.1073/pnas.1200352109
Cellulose microfibril crystallinity is reduced by mutating C-terminal transmembrane region residues CESA1A903V and CESA3T942I of cellulose synthase
Abstract
The mechanisms underlying the biosynthesis of cellulose in plants are complex and still poorly understood. A central question concerns the mechanism of microfibril structure and how this is linked to the catalytic polymerization action of cellulose synthase (CESA). Furthermore, it remains unclear whether modification of cellulose microfibril structure can be achieved genetically, which could be transformative in a bio-based economy. To explore these processes in planta, we developed a chemical genetic toolbox of pharmacological inhibitors and corresponding resistance-conferring point mutations in the C-terminal transmembrane domain region of CESA1(A903V) and CESA3(T942I) in Arabidopsis thaliana. Using (13)C solid-state nuclear magnetic resonance spectroscopy and X-ray diffraction, we show that the cellulose microfibrils displayed reduced width and an additional cellulose C4 peak indicative of a degree of crystallinity that is intermediate between the surface and interior glucans of wild type, suggesting a difference in glucan chain association during microfibril formation. Consistent with measurements of lower microfibril crystallinity, cellulose extracts from mutated CESA1(A903V) and CESA3(T942I) displayed greater saccharification efficiency than wild type. Using live-cell imaging to track fluorescently labeled CESA, we found that these mutants show increased CESA velocities in the plasma membrane, an indication of increased polymerization rate. Collectively, these data suggest that CESA1(A903V) and CESA3(T942I) have modified microfibril structure in terms of crystallinity and suggest that in plants, as in bacteria, crystallization biophysically limits polymerization.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
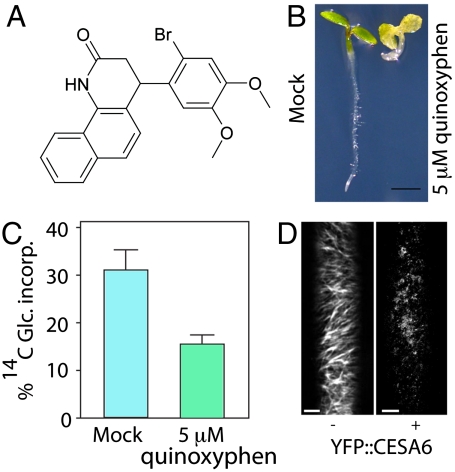
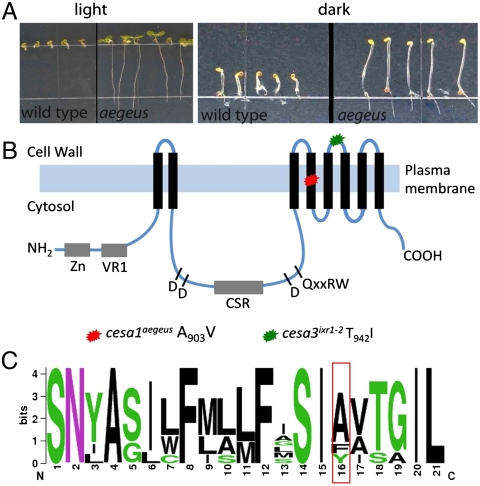
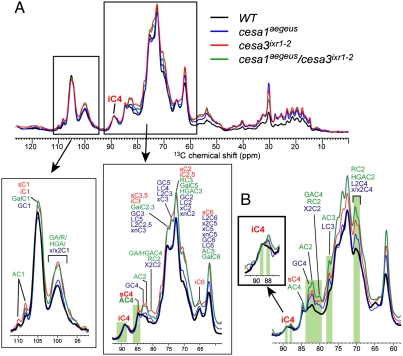
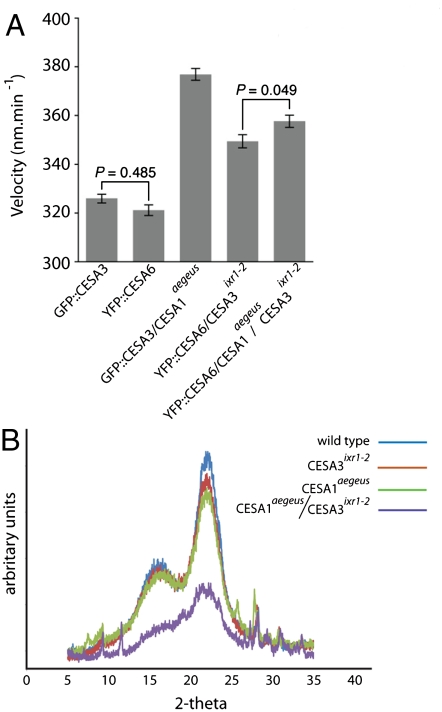
References
-
- Tsekos I. The sites of cellulose synthesis in algae: Diversity and evolution of cellulose-synthesizing enzyme complexes. J Phycol. 1999;35:635–655.
-
- Atalla RH, VanderHart DL. Native cellulose: A composite of two distinct crystalline forms. Science. 1984;223:283–285. - PubMed
-
- Nishiyama Y, Langan P, Chanzy H. Crystal structure and hydrogen-bonding system in cellulose 1b from synchrotron X-ray and neutron fiber diffraction. J Am Chem Soc. 2002;124:9074–9082. - PubMed
-
- Nishiyama Y, Sugiyama J, Chanzy H, Langan P. Crystal structure and hydrogen bonding system in cellulose 1a from synchrotron X-ray and neutron fibre diffraction. J Am Chem Soc. 2003;125:14300–14306. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
