

Insights into the phylogeny of systematically controversial haptorian ciliates (Ciliophora, Litostomatea) based on multigene analyses
- PMID: 22378805
- PMCID: PMC3350698
- DOI: 10.1098/rspb.2011.2688
Insights into the phylogeny of systematically controversial haptorian ciliates (Ciliophora, Litostomatea) based on multigene analyses
Abstract
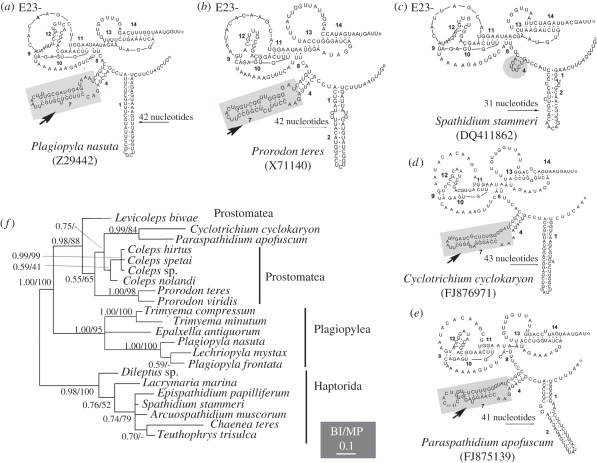
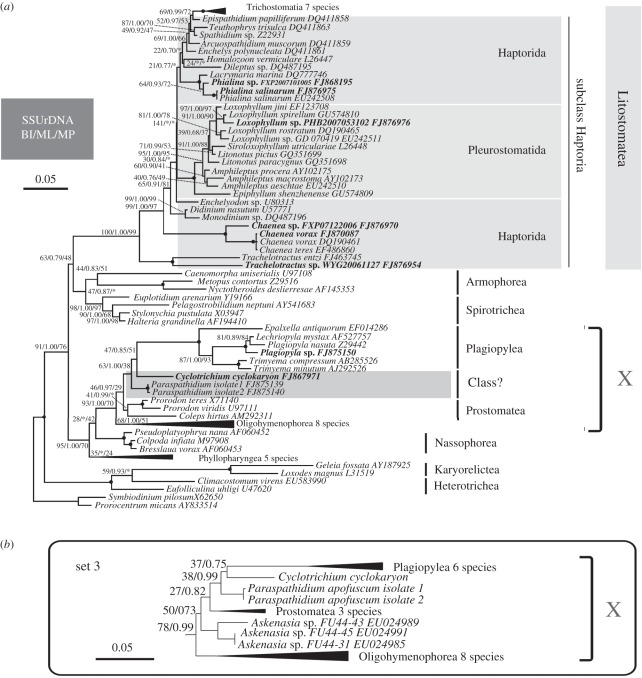
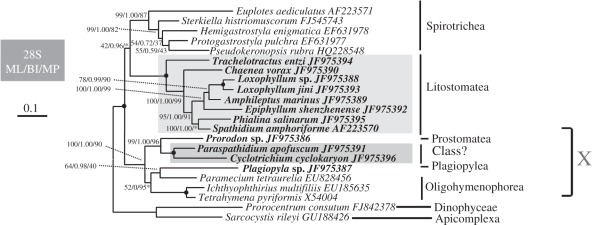
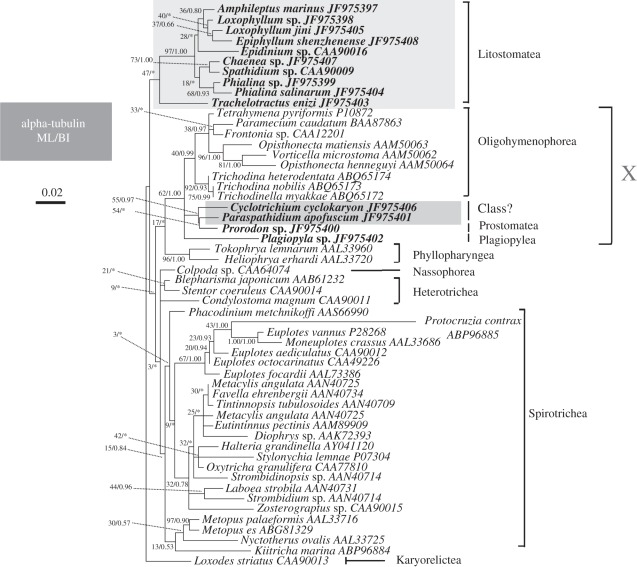
The ciliate subclass Haptoria is a diverse taxon that includes most of the free-living predators in the class Litostomatea. Phylogenetic study of this group was initially conducted using a single molecular marker small-subunit ribosomal RNA (SSU rRNA genes). Multi-gene analysis has been limited because very few other sequences were available. We performed phylogenetic analyses of Haptoria incorporating new SSU rRNA gene sequences from several debated members of the taxon, in particular, the first molecular data from Cyclotrichium. We also provided nine large-subunit ribosomal RNA (LSU rRNA) gene sequences and 10 alpha-tubulin sequences from diverse haptorians, and two possible relatives of controversial haptorians (Plagiopylea, Prostomatea). Phylogenies inferred from the different molecules showed the following: (i) Cyclotrichium and Paraspathidium were clearly separated from the haptorids and even from class Litostomatea, rejecting their high-level taxonomic assignments based on morphology. Both genera branch instead with the classes Plagiopylea, Prostomatea and Oligohymenophora. This raises the possibility that the well-known but phylogenetically problematic cyclotrichiids Mesodinium and Myrionecta may also have affinities here, rather than with litostomes; (ii) the transfer of Trachelotractus to Litostomatea is supported, especially by the analyses of SSU rRNA and LSU rRNA genes, however, Trachelotractus and Chaenea (more uncertainly) generally form the two deepest lineages within litostomes; and (iii) phylogenies of the new molecular markers are consistent with SSU rRNA gene information in recovering order Pleurostomatida as monophyletic. However, Pleurostomatida branches cladistically within order Haptorida, as does subclass Trichostomatia (on the basis of SSU rRNA phylogenies). Our results suggest that the class-level taxonomy of ciliates is still not resolved, and also that a systematic revision of litostomes is required, beginning at high taxonomic levels (taxa currently ranked as subclasses and orders).
Figures

References
-
- Lynn D. H. 2008. The ciliated protozoa: characterization, classification and guide to the literature, 3rd edn. Dordrecht, The Netherlands: Springer
-
- Foissner W., Foissner I. 1988. The fine structure of Fuscheria terricola Berger et al., 1983 and a proposed new classification of the subclass Haptoria Corliss, 1974 (Ciliophora, Litostomatea). Arch Protistenkd 135, 213–23510.1016/S0003-9365(88)80070-8 (doi:10.1016/S0003-9365(88)80070-8) - DOI - DOI
-
- Lin X., Song W., Wilbert N., Warren A. 2005. Two new marine pleurostomatid ciliates from China, Loxophyllum jini and L. qiuianum (Ciliophora, Pleurostomatida). Acta Protozool. 44, 147–157
-
- Gustafson D. E., Jr, Stoecker D. K., Johnson M. D., Van Heukelem W. F., Sneider K. 2000. Cryptophyte algae are robbed of their organelles by the marine ciliate Mesodinium rubrum. Nature 405, 1049–105210.1038/35016570 (doi:10.1038/35016570) - DOI - DOI - PubMed
-
- Hansen P. J., Fenchel T. 2006. The bloom-forming ciliate Mesodinium rubrum harbours a single permanent endosymbiont. Mar. Biol. Res. 2, 169–17710.1080/17451000600719577 (doi:10.1080/17451000600719577) - DOI - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
