

The hypersensitive sites of the murine β-globin locus control region act independently to affect nuclear localization and transcriptional elongation
- PMID: 22378846
- PMCID: PMC3335386
- DOI: 10.1182/blood-2011-09-380485
The hypersensitive sites of the murine β-globin locus control region act independently to affect nuclear localization and transcriptional elongation
Abstract
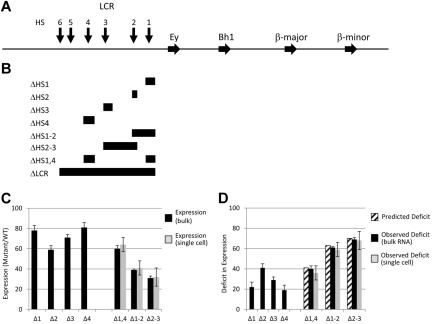
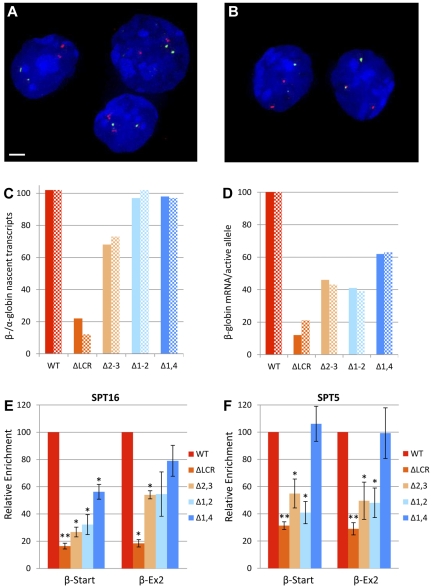
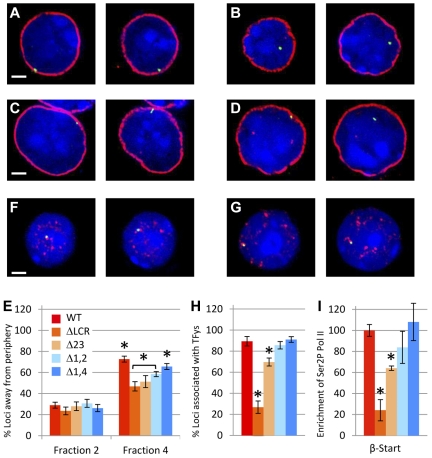
The β-globin locus control region (LCR) is necessary for high-level β-globin gene transcription and differentiation-dependent relocation of the β-globin locus from the nuclear periphery to the central nucleoplasm and to foci of hyperphosphorylated Pol II "transcription factories" (TFys). To determine the contribution of individual LCR DNaseI hypersensitive sites (HSs) to transcription and nuclear location, in the present study, we compared β-globin gene activity and location in erythroid cells derived from mice with deletions of individual HSs, deletions of 2 HSs, and deletion of the whole LCR and found all of the HSs had a similar spectrum of activities, albeit to different degrees. Each HS acts as an independent module to activate expression in an additive manner, and this is correlated with relocation away from the nuclear periphery. In contrast, HSs have redundant activities with respect to association with TFys and the probability that an allele is actively transcribed, as measured by primary RNA transcript FISH. The limiting effect on RNA levels occurs after β-globin genes associate with TFys, at which time HSs contribute to the amount of RNA arising from each burst of transcription by stimulating transcriptional elongation.
Figures
References
-
- Bender MA, Bulger M, Close J, Groudine M. Beta-globin gene switching and DNase I sensitivity of the endogenous beta-globin locus in mice do not require the locus control region. Mol Cell. 2000;5(2):387–393. - PubMed
-
- Stamatoyannopoulos G, Grosveld F. Hemoglobin switching. In: Stamatoyannopoulos G, Majerus PW, Perlmutter R, Varmus H, editors. The Molecular Basis of Blood Diseases. Philadelphia, PA: Saunders; 2001. pp. 135–182.
-
- Fraser P, Pruzina S, Antoniou M, Grosveld F. Each hypersensitive site of the human beta-globin locus control region confers a different developmental pattern of expression on the globin genes. Genes Dev. 1993;7(1):106–113. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
