

Inhibition of the NFAT pathway alleviates amyloid β neurotoxicity in a mouse model of Alzheimer's disease
- PMID: 22378890
- PMCID: PMC3296329
- DOI: 10.1523/JNEUROSCI.6439-11.2012
Inhibition of the NFAT pathway alleviates amyloid β neurotoxicity in a mouse model of Alzheimer's disease
Abstract
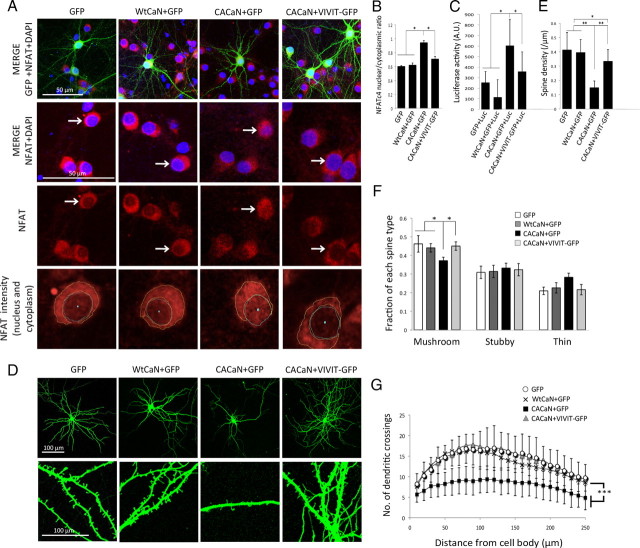
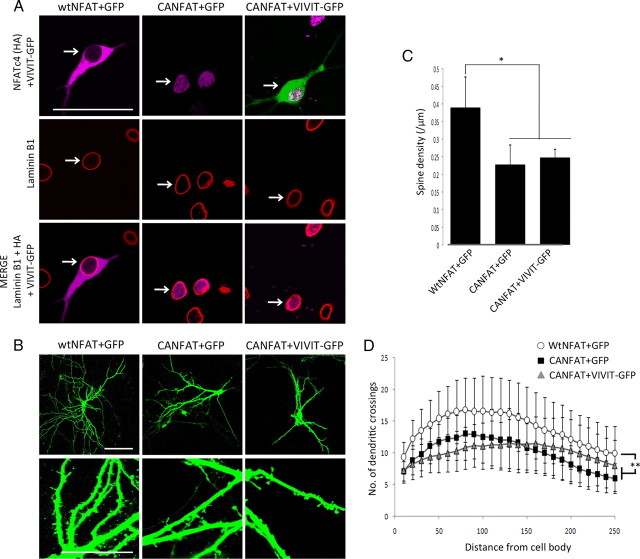
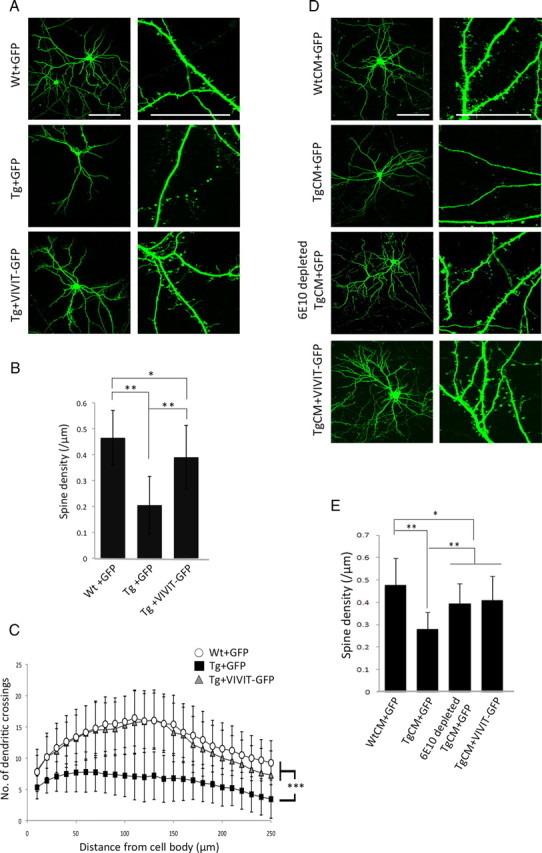
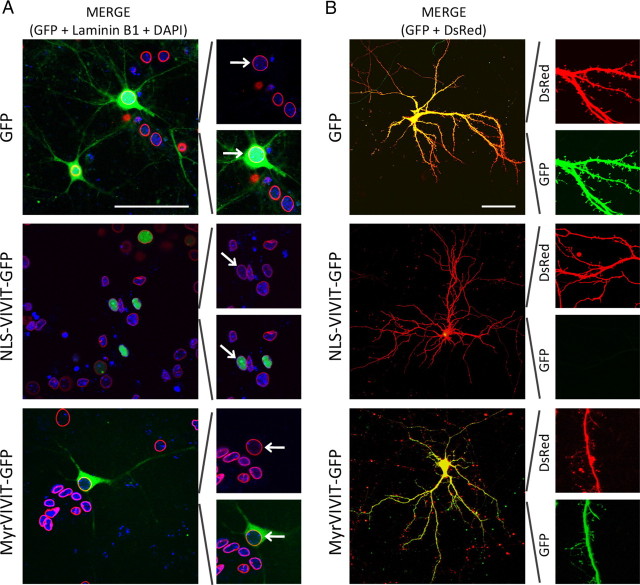
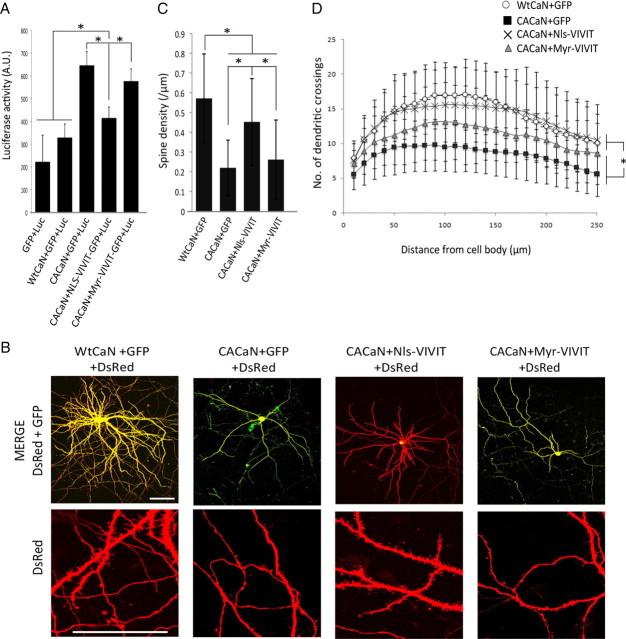
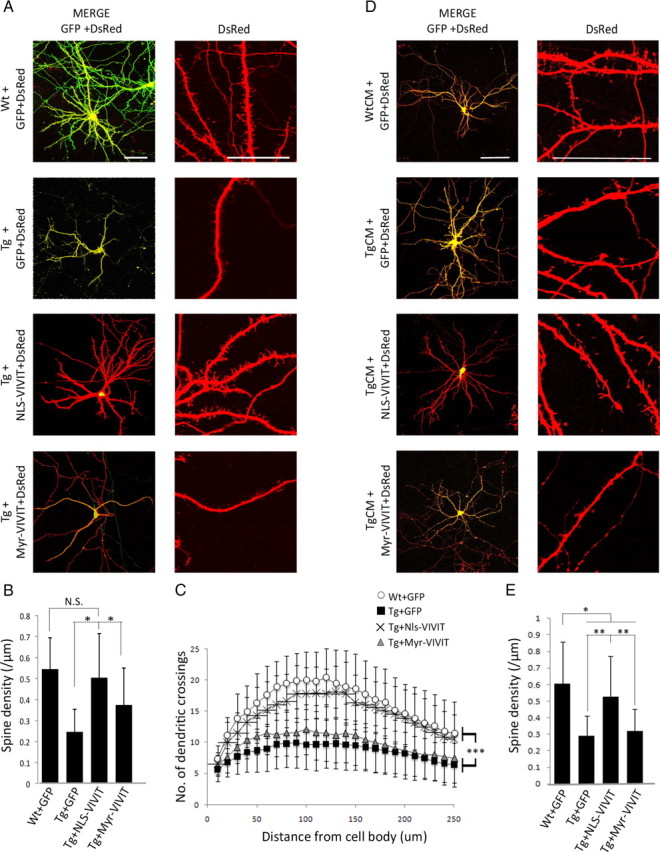
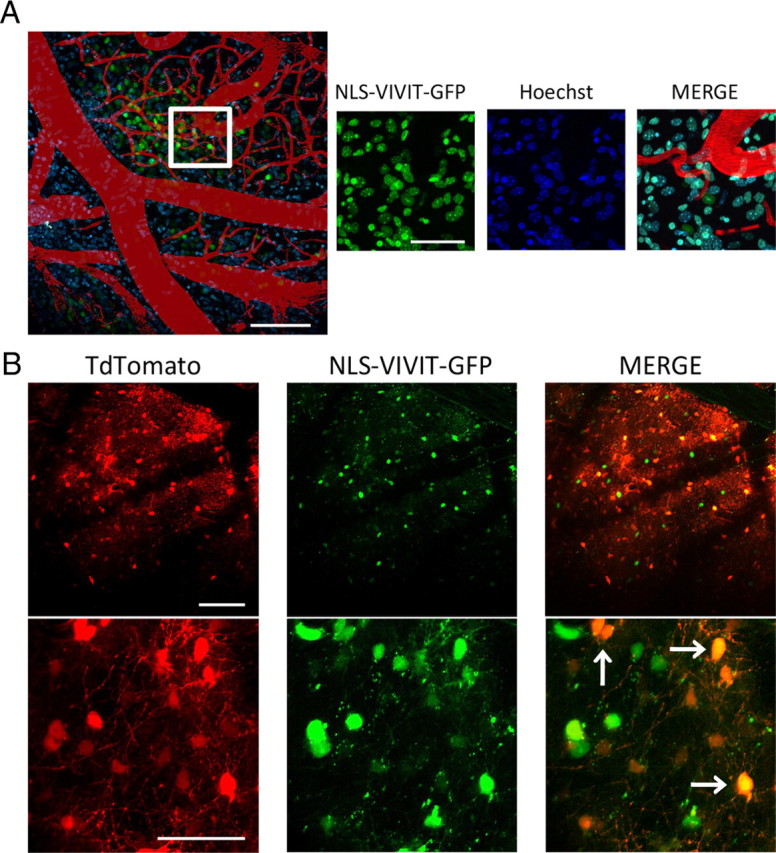
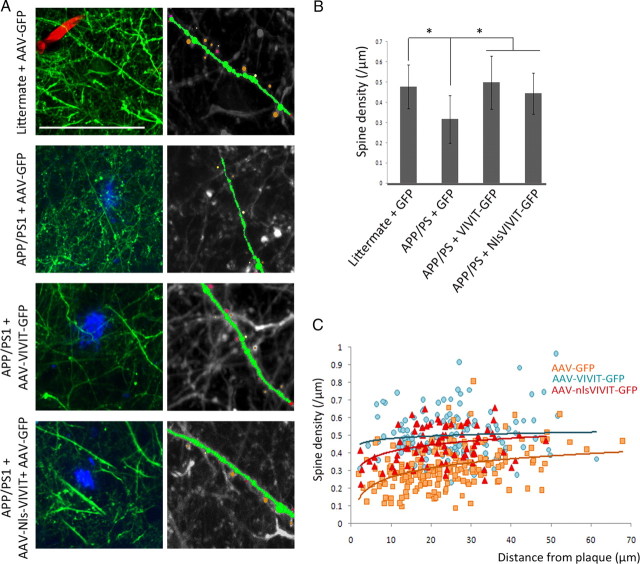
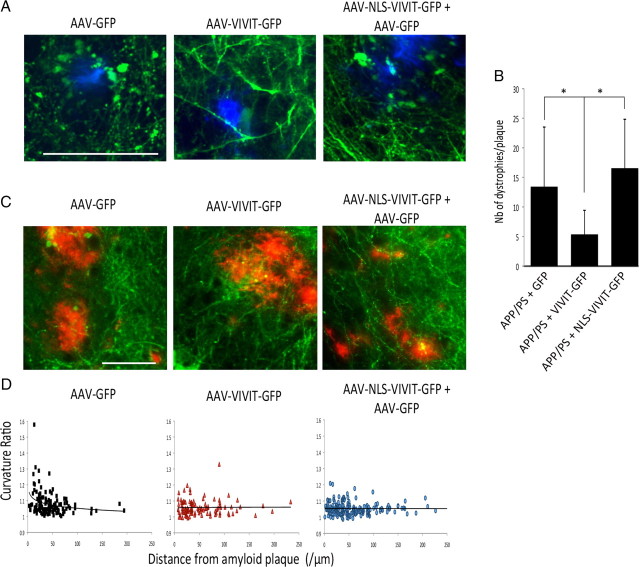
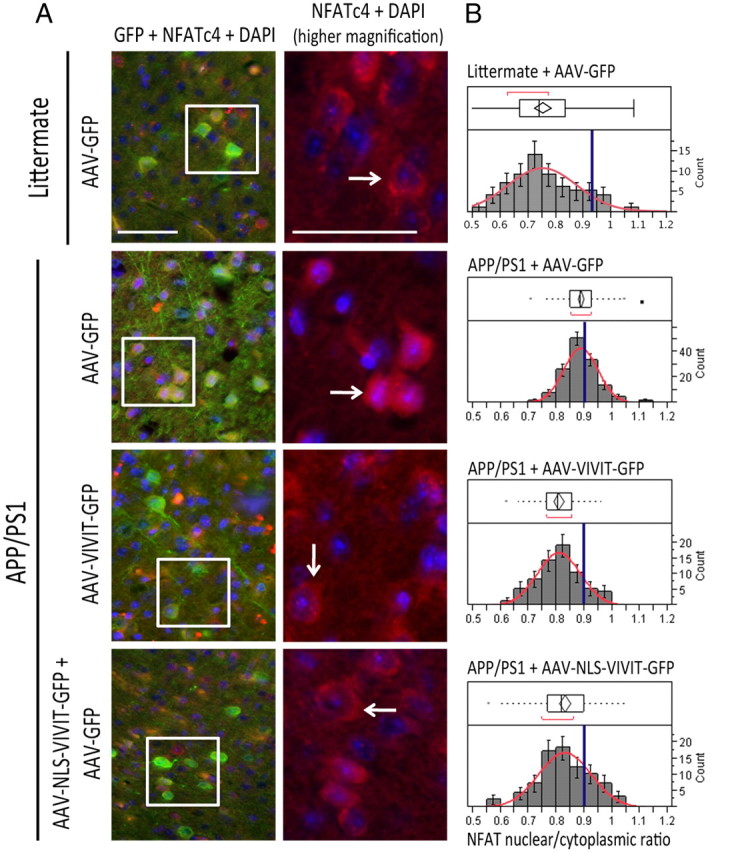
Amyloid β (Aβ) peptides, the main pathological species associated with Alzheimer's disease (AD), disturb intracellular calcium homeostasis, which in turn activates the calcium-dependent phosphatase calcineurin (CaN). CaN activation induced by Aβ leads to pathological morphological changes in neurons, and overexpression of constitutively active calcineurin is sufficient to generate a similar phenotype, even without Aβ. Here, we tested the hypothesis that calcineurin mediates neurodegenerative effects via activation of the nuclear transcription factor of activated T-cells (NFAT). We found that both spine loss and dendritic branching simplification induced by Aβ exposure were mimicked by constitutively active NFAT, and abolished when NFAT activation was blocked using the genetically encoded inhibitor VIVIT. When VIVIT was specifically addressed to the nucleus, identical beneficial effects were observed, thus enforcing the role of NFAT transcriptional activity in Aβ-related neurotoxicity. In vivo, when VIVIT or its nuclear counterpart were overexpressed in a transgenic model of Alzheimer's disease via a gene therapy approach, the spine loss and neuritic abnormalities observed in the vicinity of amyloid plaques were blocked. Overall, these results suggest that NFAT/calcineurin transcriptional cascades contribute to Aβ synaptotoxicity, and may provide a new specific set of pathways for neuroprotective strategies.
Figures
Comment in
-
The Aβ-induced NFAT apoptotic pathway is also activated by GSK-3 inhibition: implications for Alzheimer therapeutics.J Neurosci. 2012 Jul 11;32(28):9454-6. doi: 10.1523/JNEUROSCI.2143-12.2012. J Neurosci. 2012. PMID: 22787030 Free PMC article. No abstract available.
References
-
- Amberg GC, Rossow CF, Navedo MF, Santana LF. NFATc3 regulates Kv2.1 expression in arterial smooth muscle. J Biol Chem. 2004;279:47326–47334. - PubMed
-
- Aramburu J, Yaffe MB, López-Rodríguez C, Cantley LC, Hogan PG, Rao A. Affinity-driven peptide selection of an NFAT inhibitor more selective than cyclosporin A. Science. 1999;285:2129–2133. - PubMed
-
- Bacskai BJ, Klunk WE, Mathis CA, Hyman BT. Imaging amyloid-beta deposits in vivo. J Cereb Blood Flow Metab. 2002;22:1035–1041. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases