

Suppression of the epithelial-mesenchymal transition by Grainyhead-like-2
- PMID: 22379025
- PMCID: PMC3342427
- DOI: 10.1158/0008-5472.CAN-11-4038
Suppression of the epithelial-mesenchymal transition by Grainyhead-like-2
Abstract
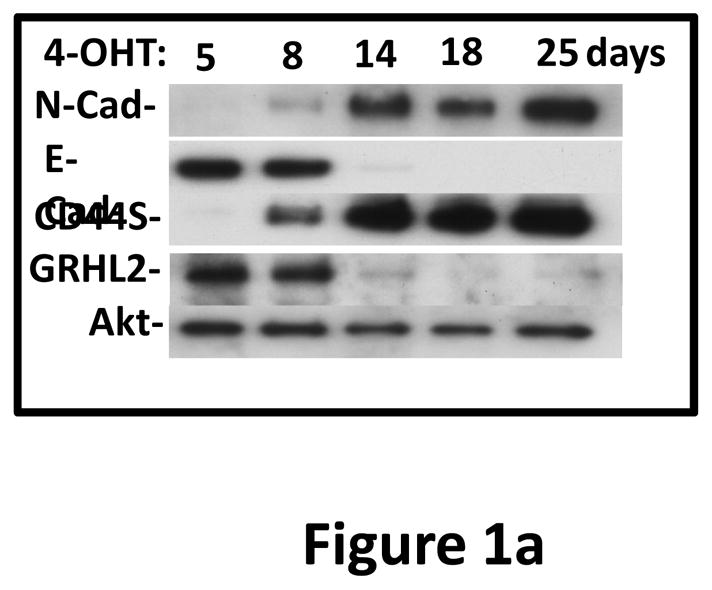
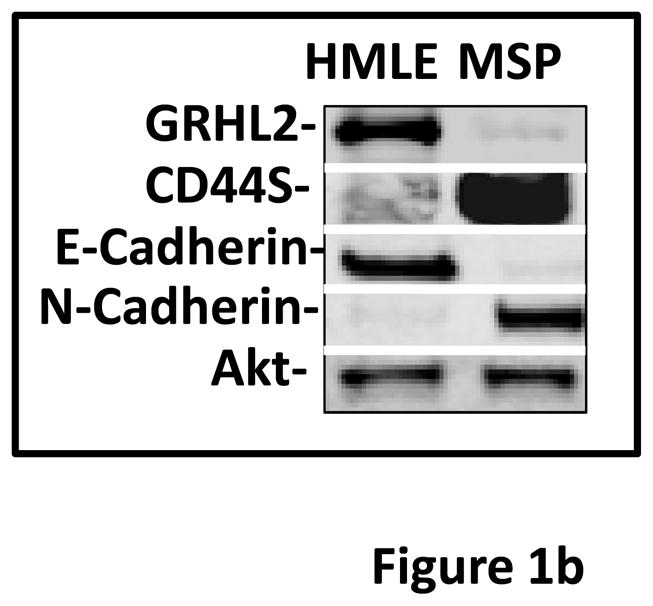
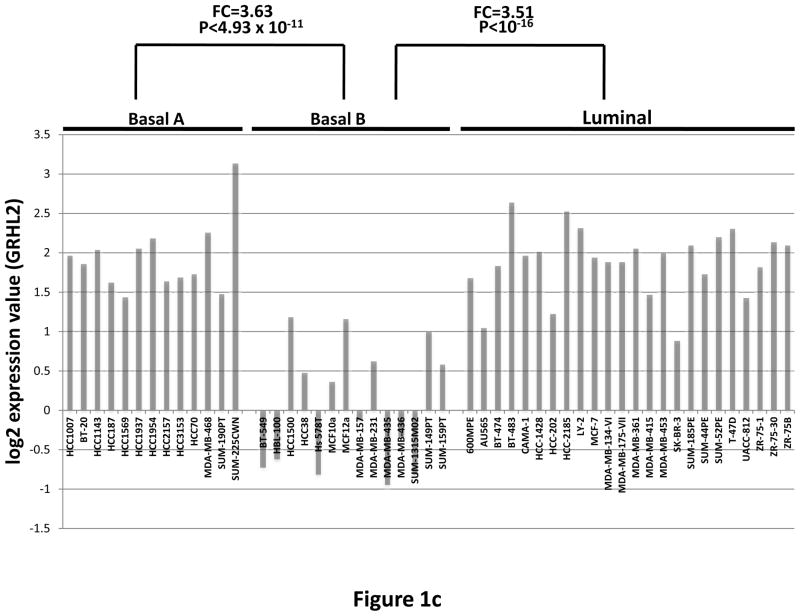
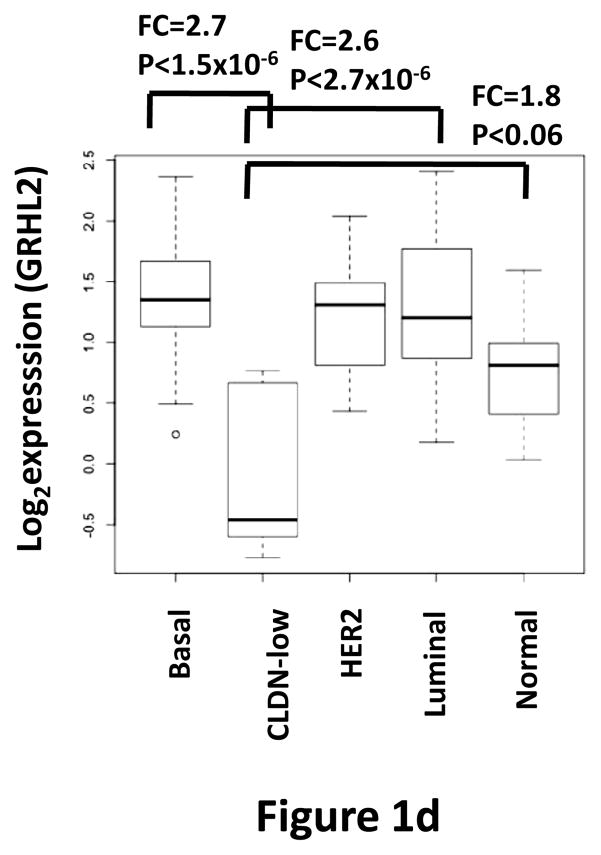
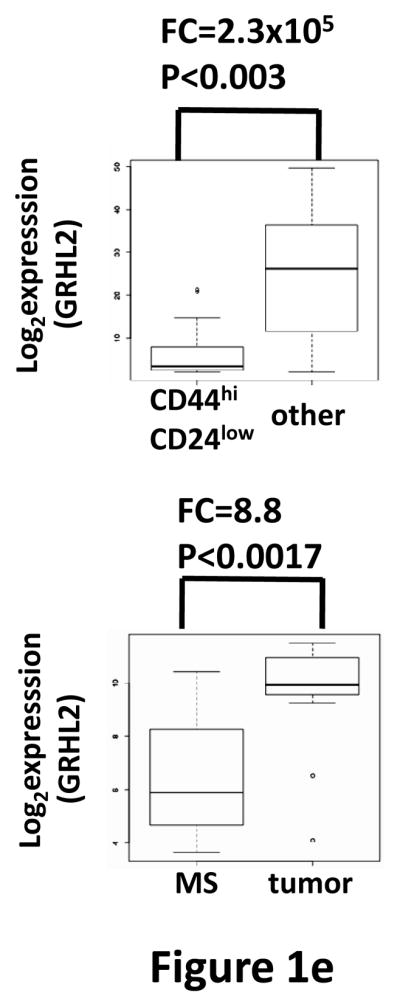
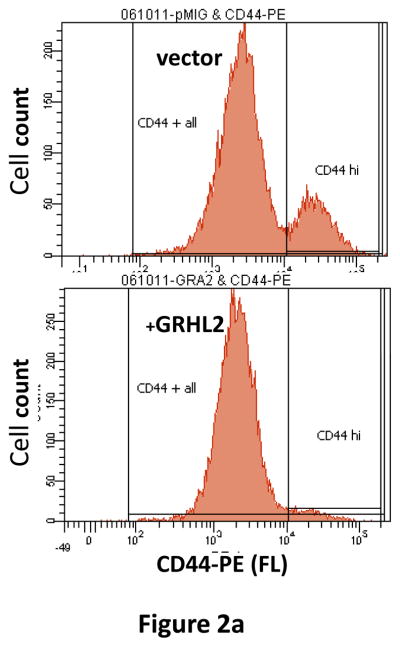
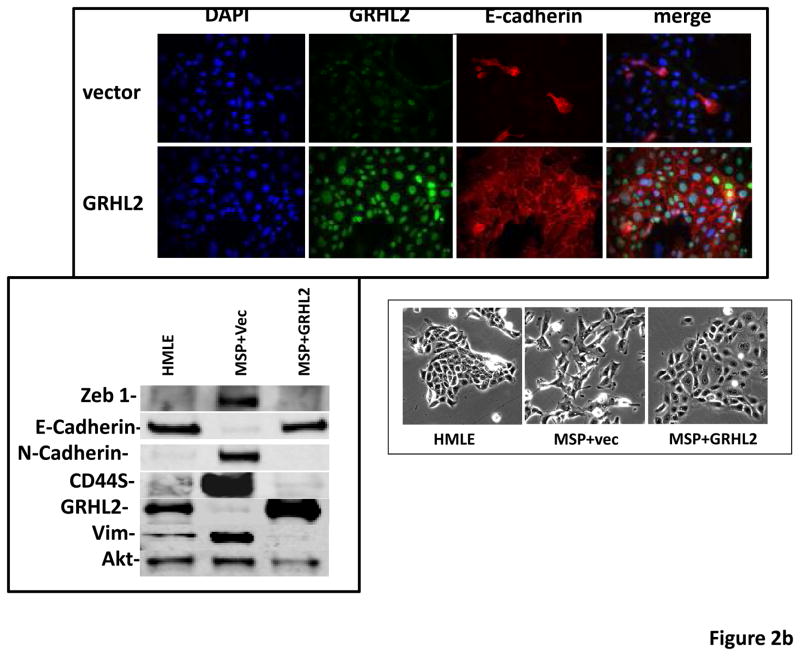
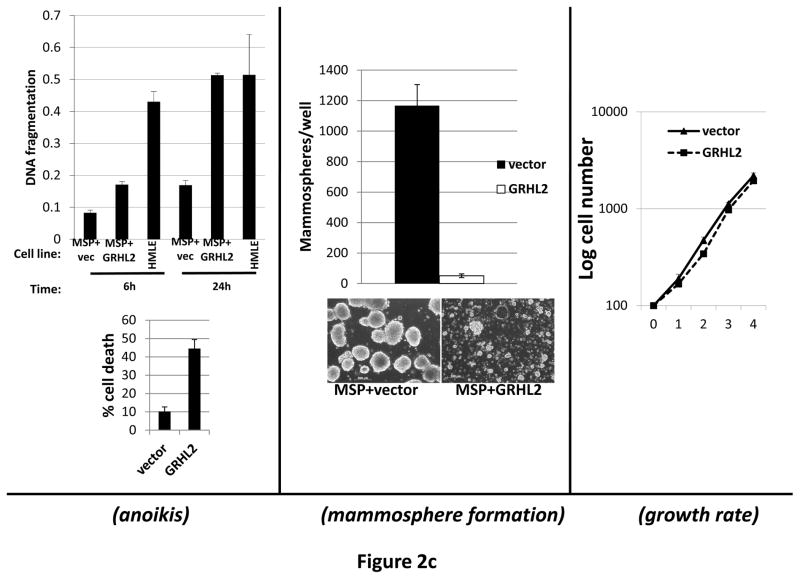
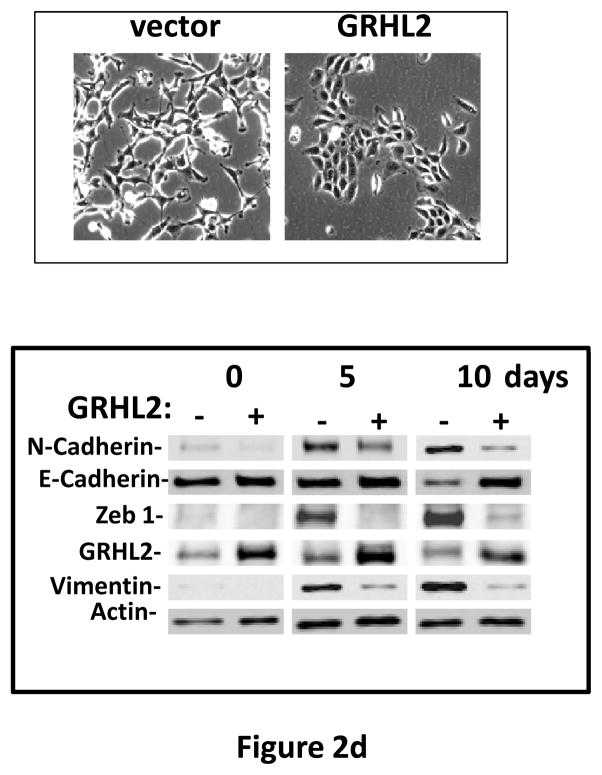
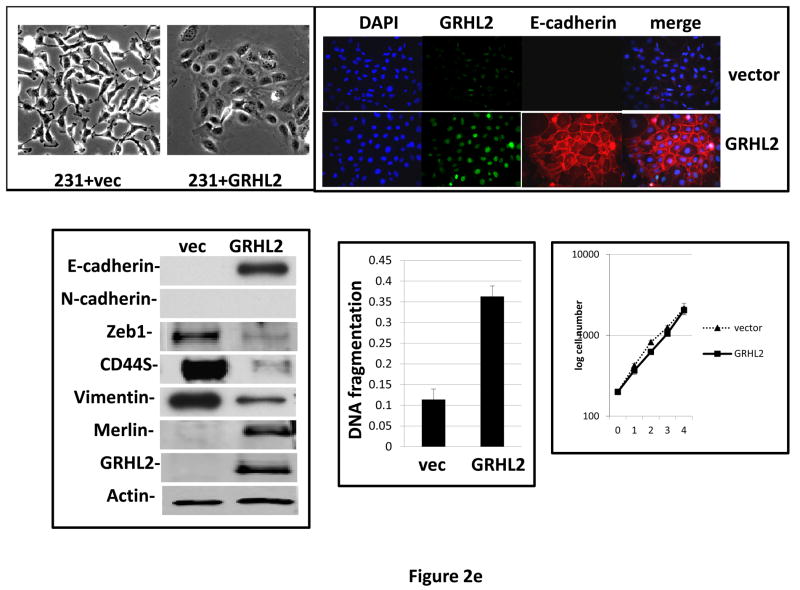
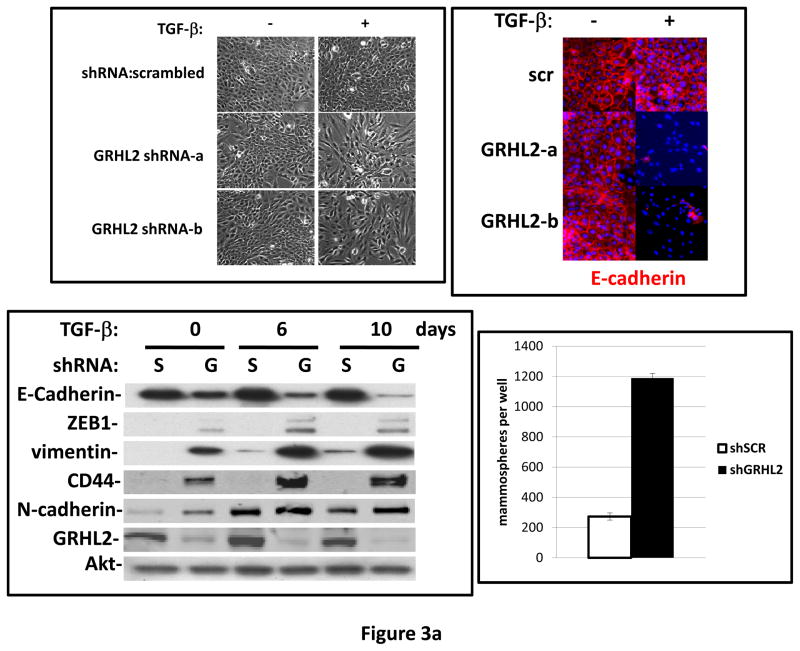
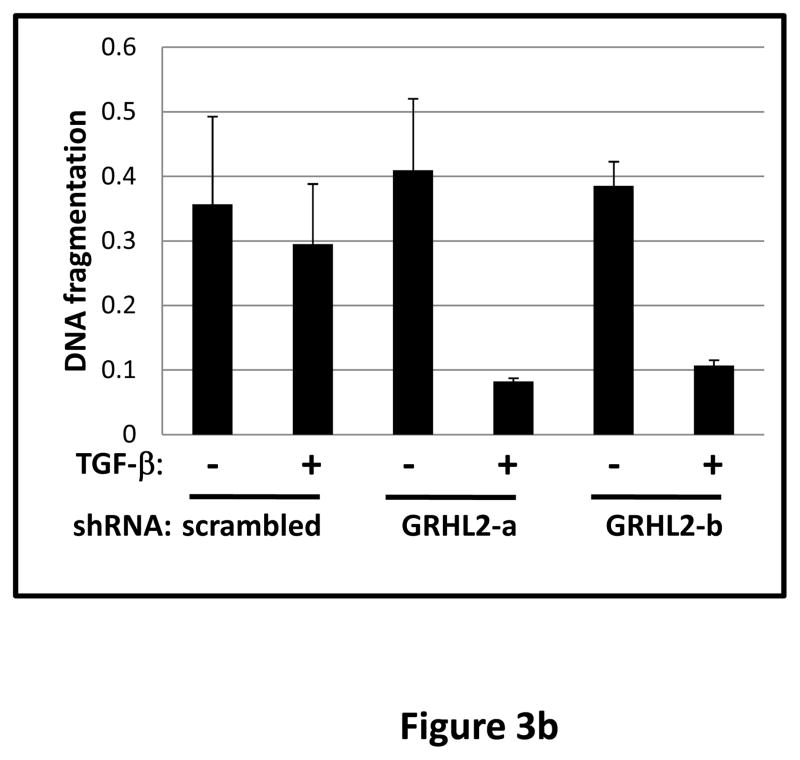
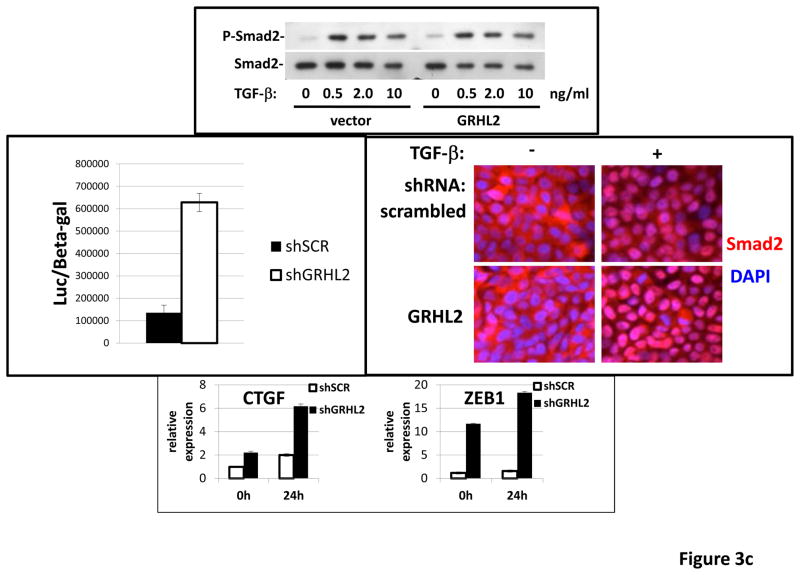
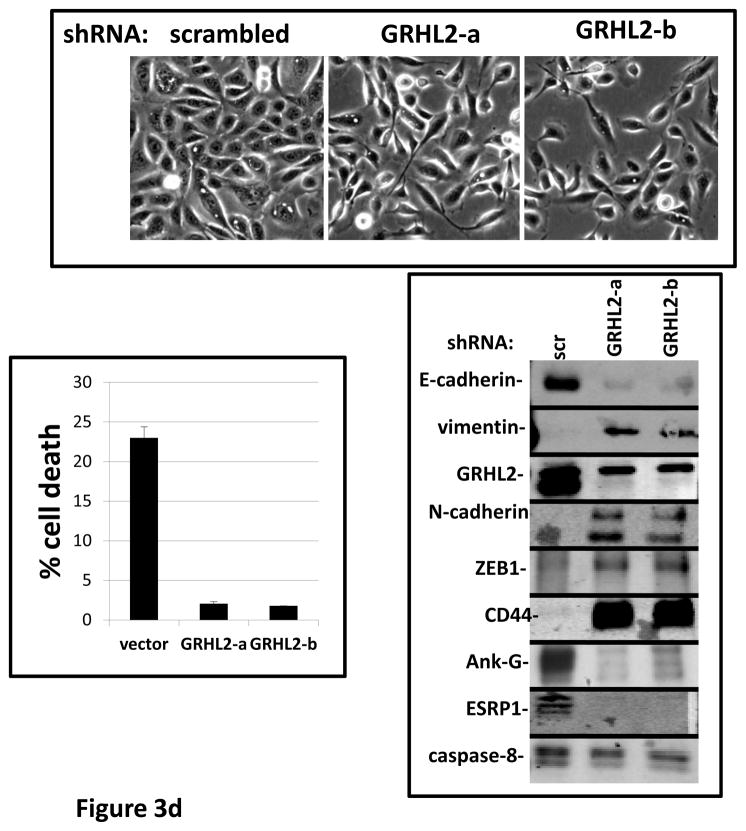
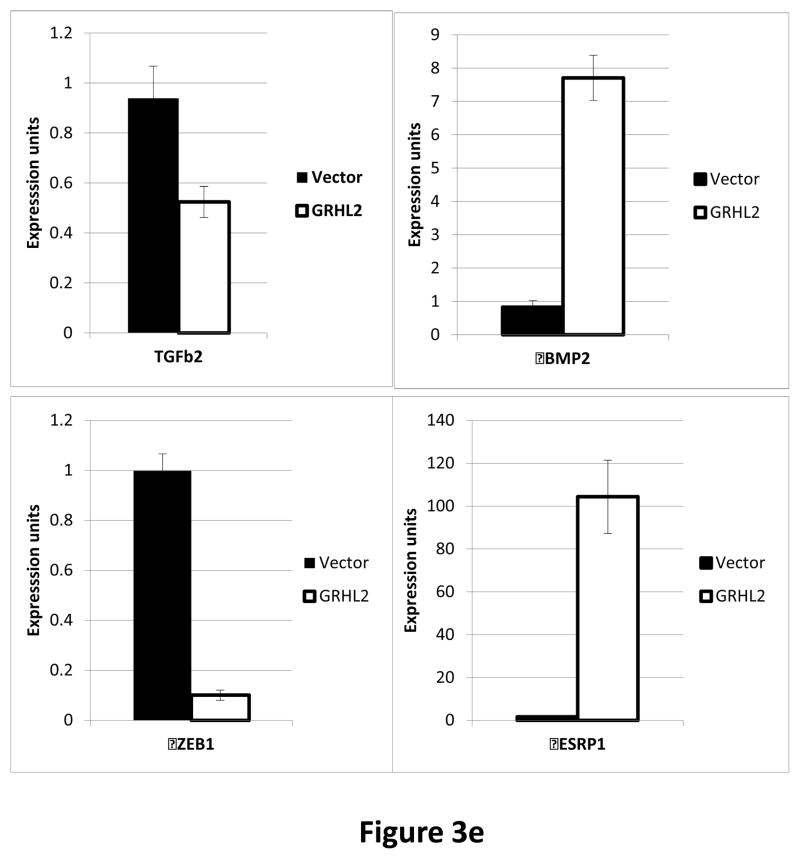
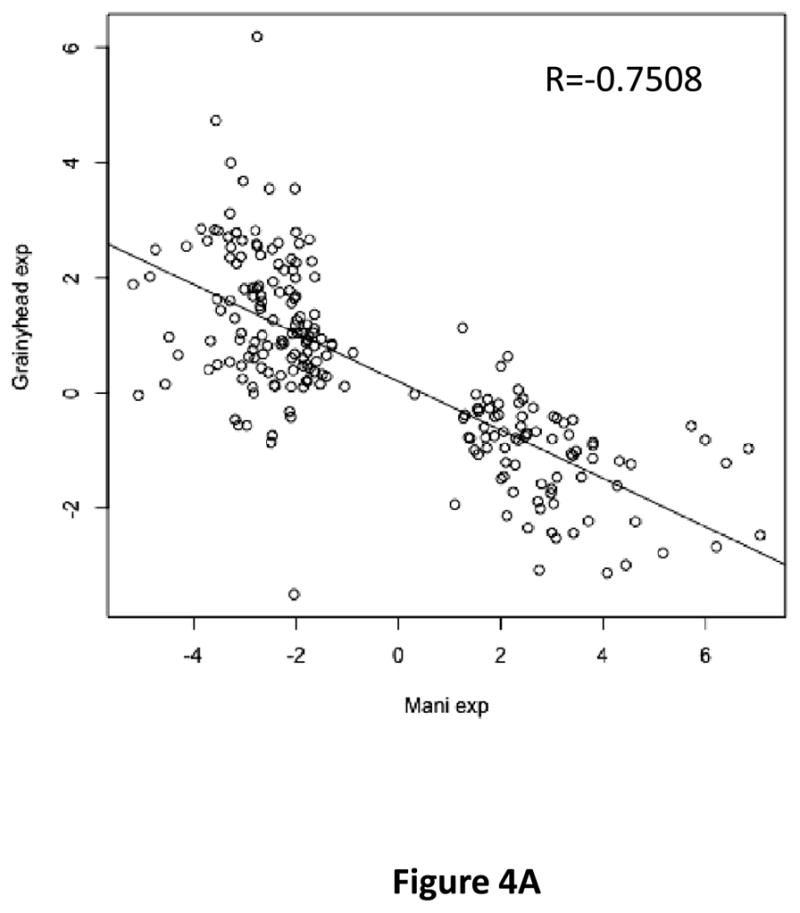
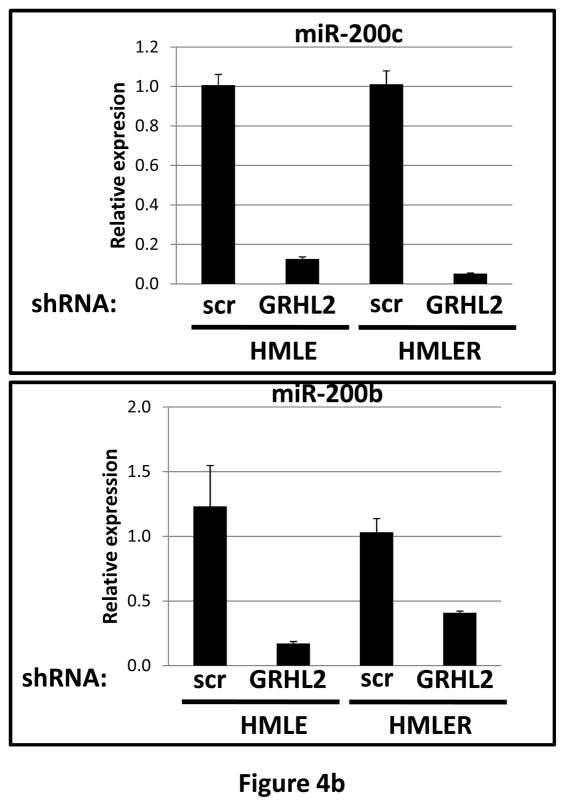
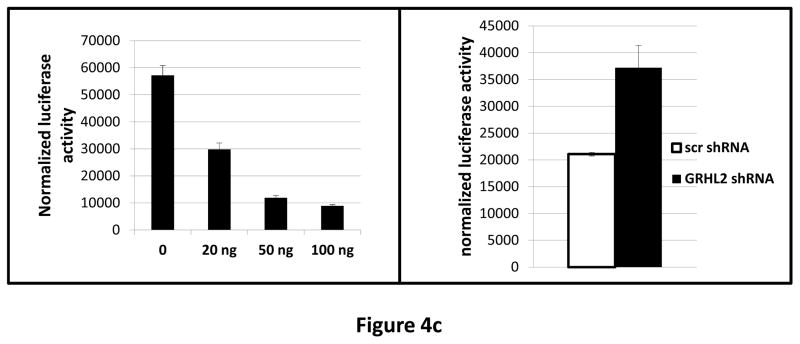
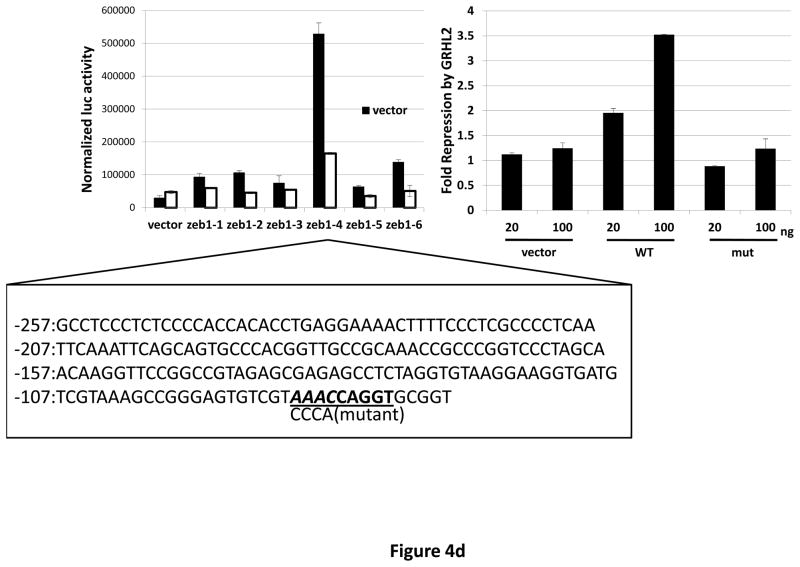
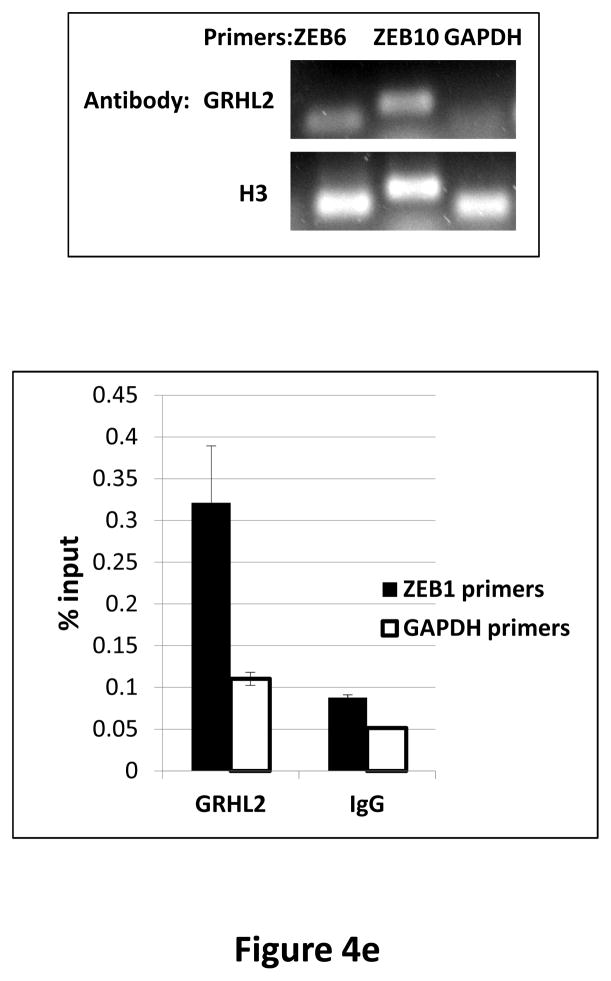
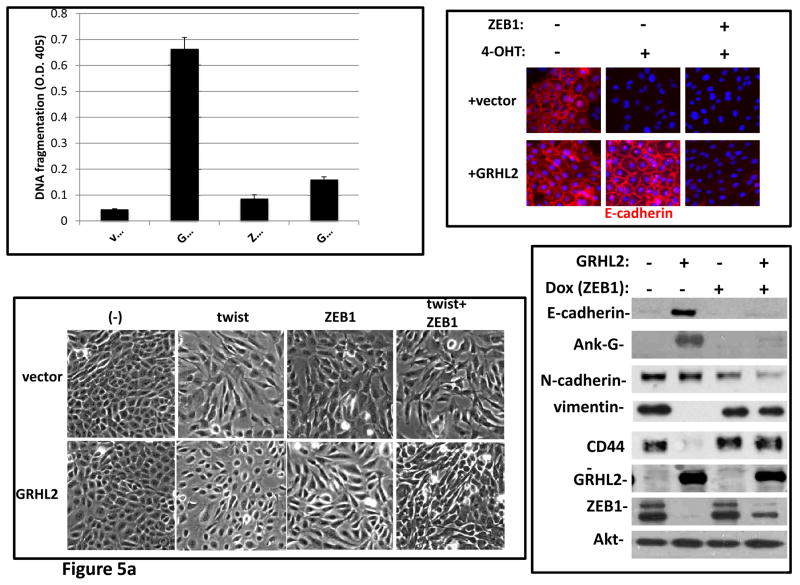
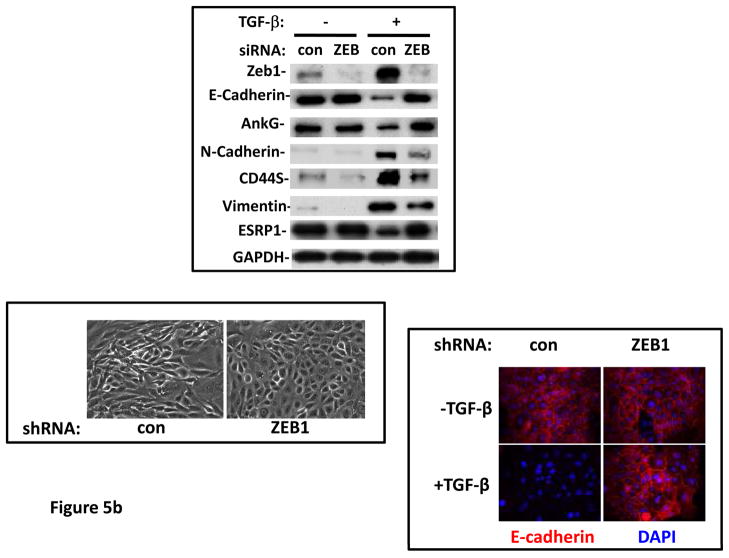
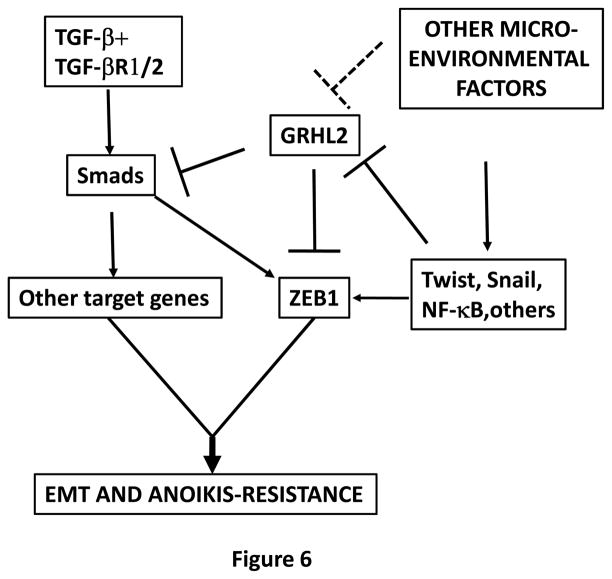
Grainyhead genes are involved in wound healing and developmental neural tube closure. In light of the high degree of similarity between the epithelial-mesenchymal transitions (EMT) occurring in wound-healing processes and the cancer stem cell-like compartment of tumors, including TGF-β dependence, we investigated the role of the Grainyhead gene, Grainyhead-like-2 (GRHL2) in oncogenic EMT. GRHL2 was downregulated specifically in the claudin-low subclass breast tumors and in basal-B subclass breast cancer cell lines. GRHL2 suppressed TGF-β-induced, Twist-induced or spontaneous EMT, enhanced anoikis sensitivity, and suppressed mammosphere generation in mammary epithelial cells. These effects were mediated in part by suppression of ZEB1 expression via direct repression of the ZEB1 promoter. GRHL2 also inhibited Smad-mediated transcription and it upregulated mir-200b/c as well as the TGF-β receptor antagonist, BMP2. Finally, ectopic expression of GRHL2 in MDA-MB-231 breast cancer cells triggered an MET and restored sensitivity to anoikis. Taken together, our findings define a major role for GRHL2 in the suppression of oncogenic EMT in breast cancer cells.
©2012 AACR
Figures
References
-
- Polyak K, Weinberg RA. Transitions between epithelial and mesenchymal states: acquisition of malignant and stem cell traits. Nat Rev Cancer. 2009;9:265–73. - PubMed
-
- Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009;139:871–90. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
