

The contribution of systemic and pulmonary immune effectors to vaccine-induced protection from H5N1 influenza virus infection
- PMID: 22379093
- PMCID: PMC3347357
- DOI: 10.1128/JVI.07205-11
The contribution of systemic and pulmonary immune effectors to vaccine-induced protection from H5N1 influenza virus infection
Abstract
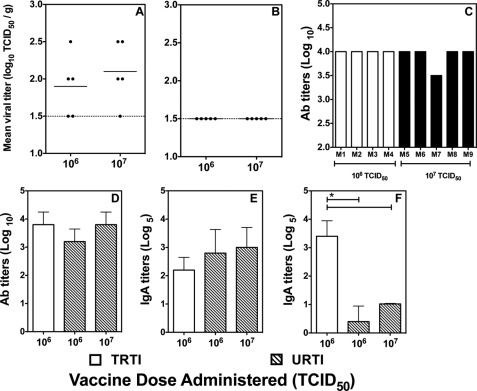
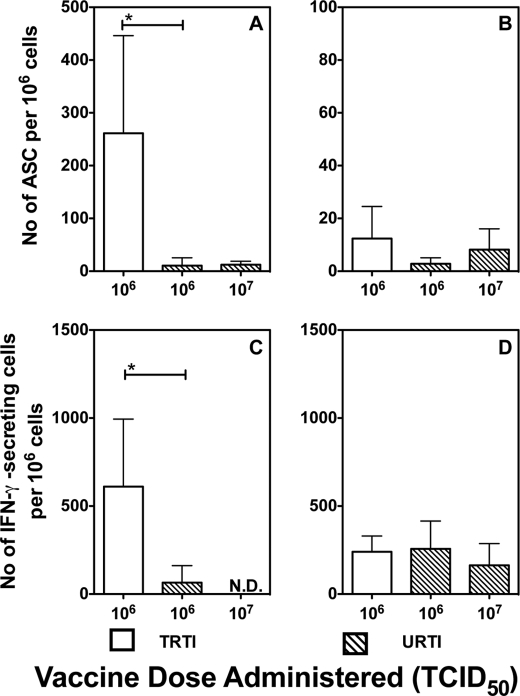
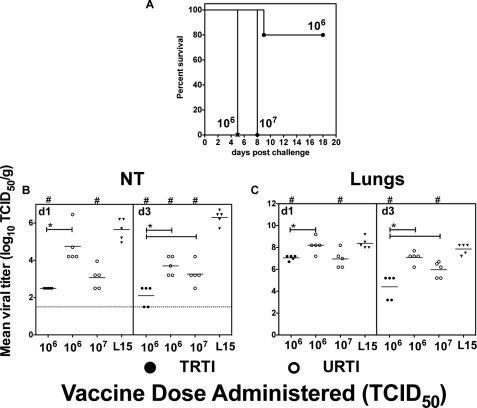
Live attenuated influenza vaccines (LAIVs) are effective in providing protection against influenza challenge in animal models and in preventing disease in humans. We previously showed that LAIVs elicit a range of immune effectors and that successful induction of pulmonary cellular and humoral immunity in mice requires pulmonary replication of the vaccine virus. An upper respiratory tract immunization (URTI) model was developed in mice to mimic the human situation, in which the vaccine virus does not replicate in the lower respiratory tract, allowing us to assess the protective efficacy of an H5N1 LAIV against highly pathogenic H5N1 virus challenge in the absence of significant pulmonary immunity. Our results show that, after one dose of an H5N1 LAIV, pulmonary influenza-specific lymphocytes are the main contributors to clearance of challenge virus from the lungs and that contributions of influenza-specific enzyme-linked immunosorbent assay (ELISA) antibodies in serum and splenic CD8(+) T cells were negligible. Complete protection from H5N1 challenge was achieved after two doses of H5N1 LAIV and was associated with maturation of the antibody response. Although passive transfer of sera from mice that received two doses of vaccine prevented lethality in naive recipients following challenge, the mice showed significant weight loss, with high pulmonary titers of the H5N1 virus. These data highlight the importance of mucosal immunity in mediating optimal protection against H5N1 infection. Understanding the requirements for effective induction and establishment of these protective immune effectors in the respiratory tract paves the way for a more rational and effective vaccine approach in the future.
Figures
Similar articles
-
The magnitude of local immunity in the lungs of mice induced by live attenuated influenza vaccines is determined by local viral replication and induction of cytokines.J Virol. 2011 Jan;85(1):76-85. doi: 10.1128/JVI.01564-10. Epub 2010 Oct 20. J Virol. 2011. PMID: 20962087 Free PMC article.
-
M2SR, a novel live influenza vaccine, protects mice and ferrets against highly pathogenic avian influenza.Vaccine. 2017 Jul 24;35(33):4177-4183. doi: 10.1016/j.vaccine.2017.06.039. Epub 2017 Jun 28. Vaccine. 2017. PMID: 28668565 Free PMC article.
-
Mosaic H5 Hemagglutinin Provides Broad Humoral and Cellular Immune Responses against Influenza Viruses.J Virol. 2016 Jul 11;90(15):6771-6783. doi: 10.1128/JVI.00730-16. Print 2016 Aug 1. J Virol. 2016. PMID: 27194759 Free PMC article.
-
Immune responses to infection with H5N1 influenza virus.Virus Res. 2013 Dec 5;178(1):44-52. doi: 10.1016/j.virusres.2013.05.011. Epub 2013 Jun 2. Virus Res. 2013. PMID: 23735534 Free PMC article. Review.
-
The influence of pregnancy on systemic immunity.Immunol Res. 2012 Dec;54(1-3):254-61. doi: 10.1007/s12026-012-8303-9. Immunol Res. 2012. PMID: 22447351 Free PMC article. Review.
Cited by
-
Polymeric Nanoparticle-Based Vaccine Adjuvants and Delivery Vehicles.Curr Top Microbiol Immunol. 2021;433:29-76. doi: 10.1007/82_2020_226. Curr Top Microbiol Immunol. 2021. PMID: 33165869 Free PMC article. Review.
-
Intranodal administration of mRNA encoding nucleoprotein provides cross-strain immunity against influenza in mice.J Transl Med. 2019 Jul 25;17(1):242. doi: 10.1186/s12967-019-1991-3. J Transl Med. 2019. PMID: 31345237 Free PMC article.
-
Pseudotyped influenza A virus as a vaccine for the induction of heterotypic immunity.J Virol. 2012 Dec;86(24):13397-406. doi: 10.1128/JVI.01820-12. Epub 2012 Sep 26. J Virol. 2012. PMID: 23015719 Free PMC article.
-
Simultaneous Aerosol and Intramuscular Immunization with Influenza Vaccine Induces Powerful Protective Local T Cell and Systemic Antibody Immune Responses in Pigs.J Immunol. 2021 Feb 1;206(3):652-663. doi: 10.4049/jimmunol.2001086. Epub 2020 Dec 16. J Immunol. 2021. PMID: 33328212 Free PMC article.
-
Aerosol immunization with influenza matrix, nucleoprotein, or both prevents lung disease in pig.NPJ Vaccines. 2024 Oct 13;9(1):188. doi: 10.1038/s41541-024-00989-8. NPJ Vaccines. 2024. PMID: 39397062 Free PMC article.
References
-
- Belshe R, et al. 2000. Correlates of immune protection induced by live, attenuated, cold-adapted, trivalent, intranasal influenza virus vaccine. J. Infect. Dis. 181:1133–1137 - PubMed
-
- Belshe RB, et al. 1998. The efficacy of live attenuated, cold-adapted, trivalent, intranasal influenzavirus vaccine in children. N. Engl. J. Med. 338:1405–1412 - PubMed
-
- Bernstein DI, et al. 2003. Effect of yearly vaccinations with live, attenuated, cold-adapted, trivalent, intranasal influenza vaccines on antibody responses in children. Pediatr. Infect. Dis. J. 22:28–34 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
