

Inositol 5-phosphatases: insights from the Lowe syndrome protein OCRL
- PMID: 22381590
- PMCID: PMC3323734
- DOI: 10.1016/j.tibs.2012.01.002
Inositol 5-phosphatases: insights from the Lowe syndrome protein OCRL
Abstract
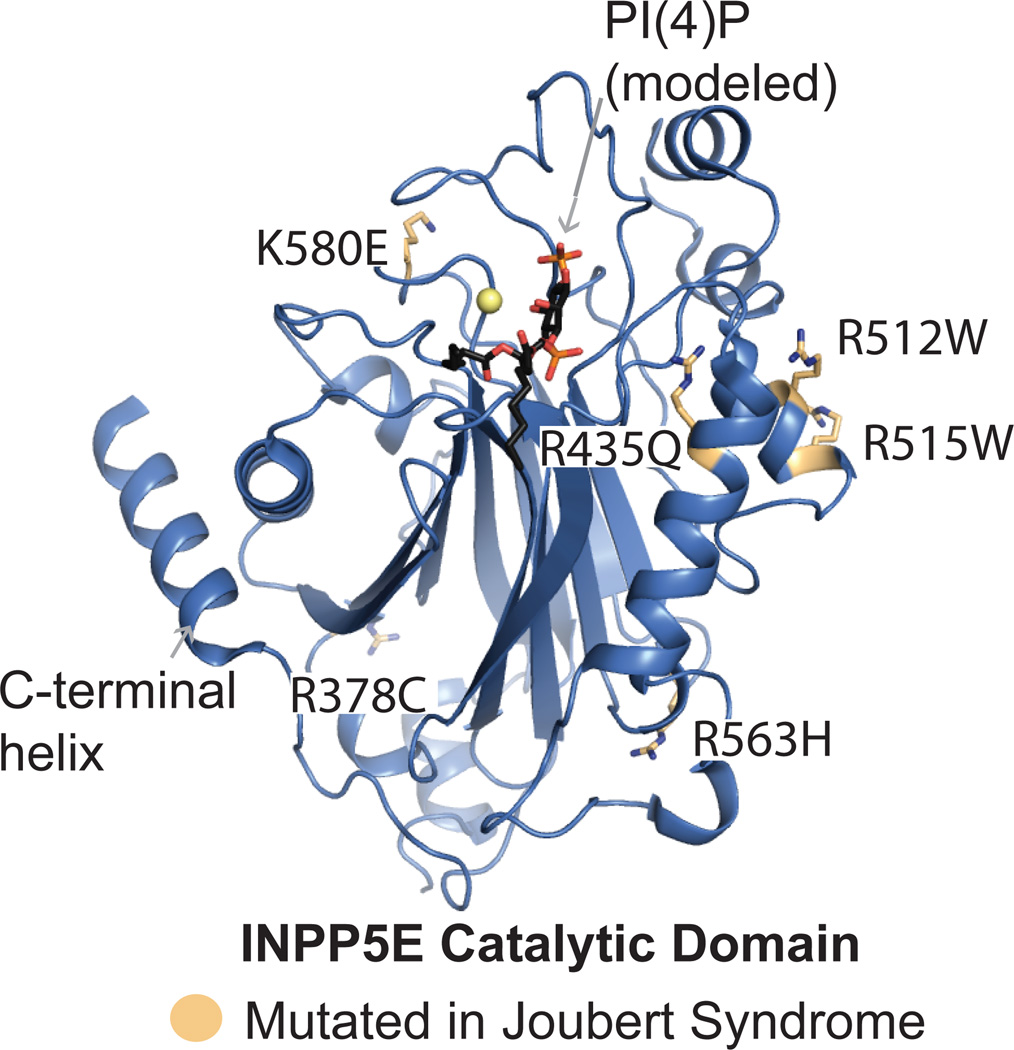
The precise regulation of phosphoinositide lipids in cellular membranes is crucial for cellular survival and function. Inositol 5-phosphatases have been implicated in a variety of disorders, including various cancers, obesity, type 2 diabetes, neurodegenerative diseases and rare genetic conditions. Despite the obvious impact on human health, relatively little structural and biochemical information is available for this family. Here, we review recent structural and mechanistic work on the 5-phosphatases with a focus on OCRL, whose loss of function results in oculocerebrorenal syndrome of Lowe and Dent 2 disease. Studies of OCRL emphasize how the actions of 5-phosphatases rely on both intrinsic and extrinsic membrane recognition properties for full catalytic function. Additionally, structural analysis of missense mutations in the catalytic domain of OCRL provides insight into the phenotypic heterogeneity observed in Lowe syndrome and Dent disease.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
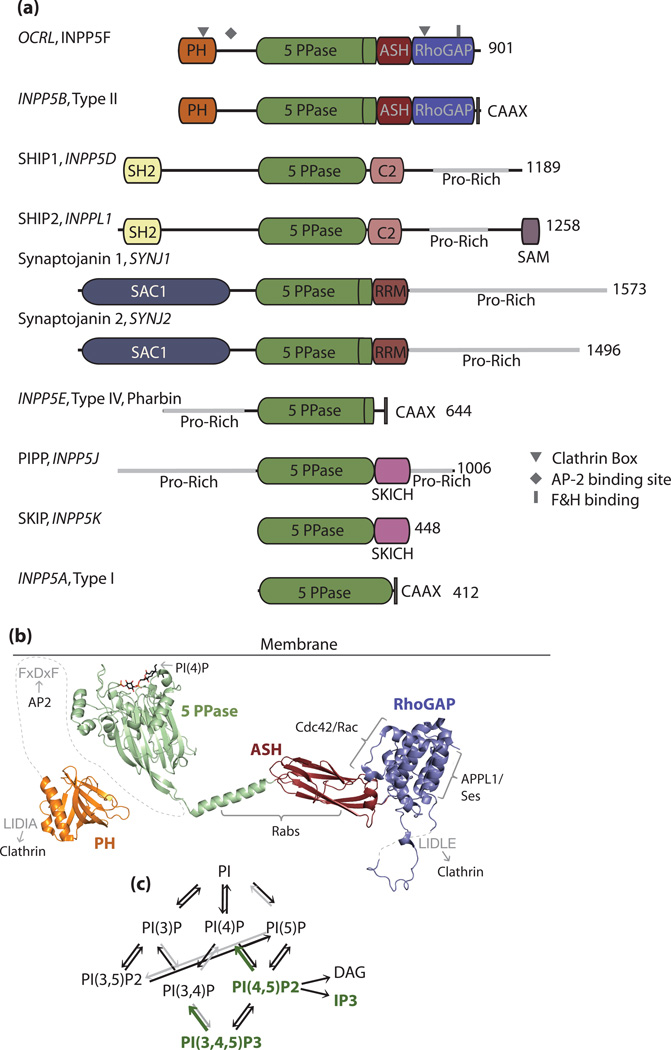
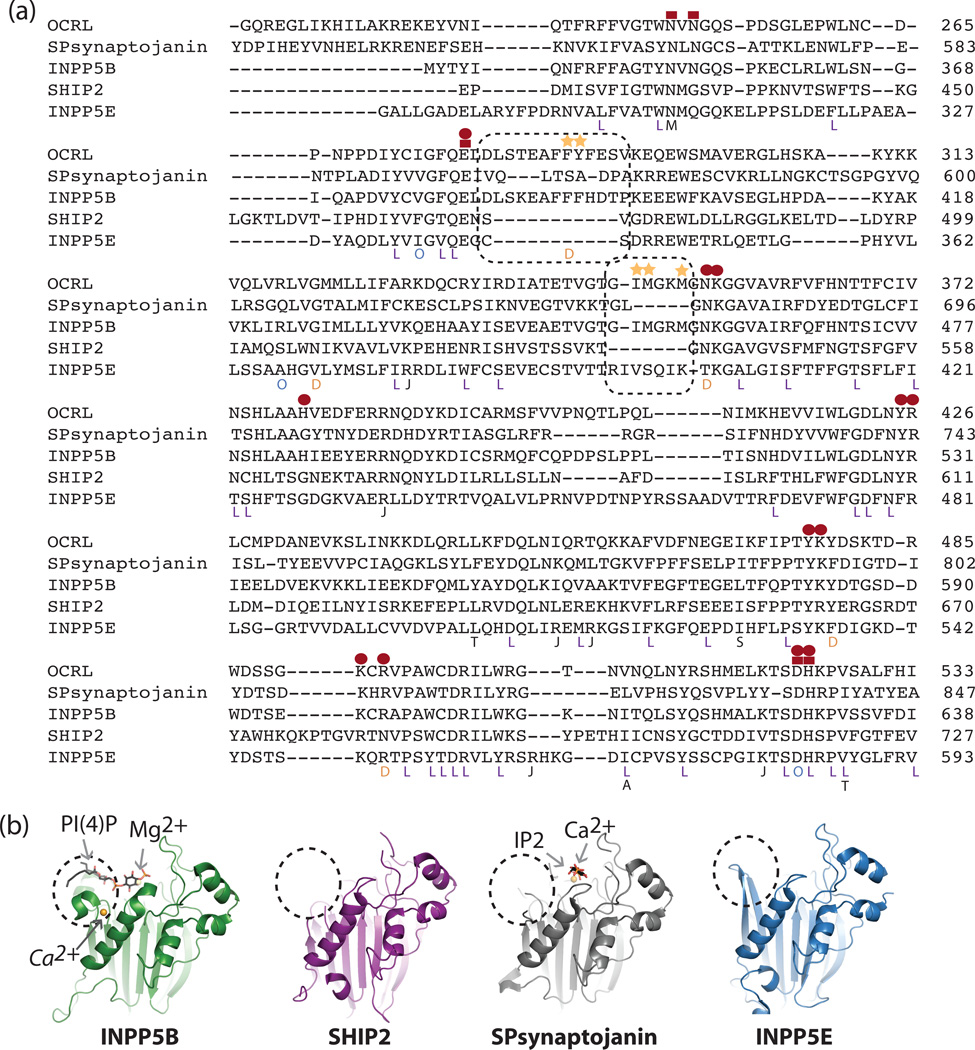
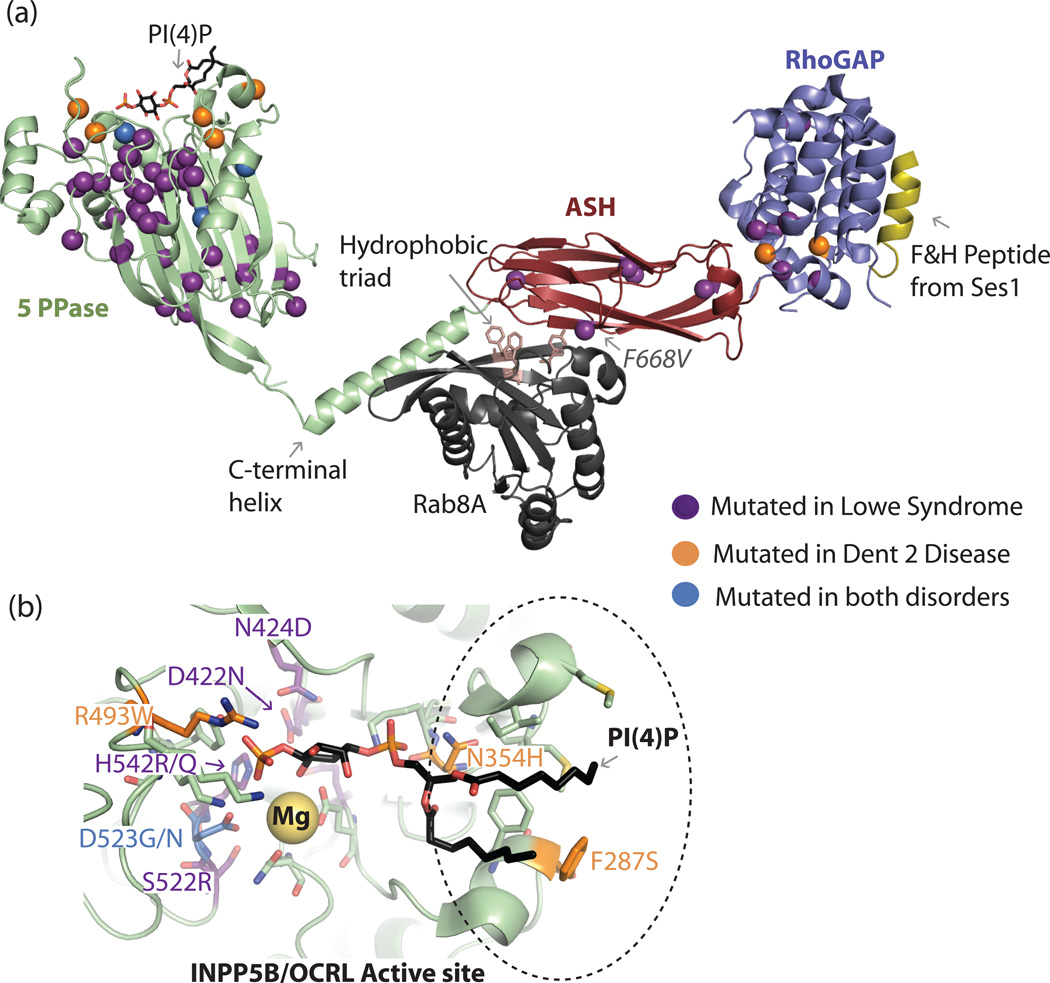
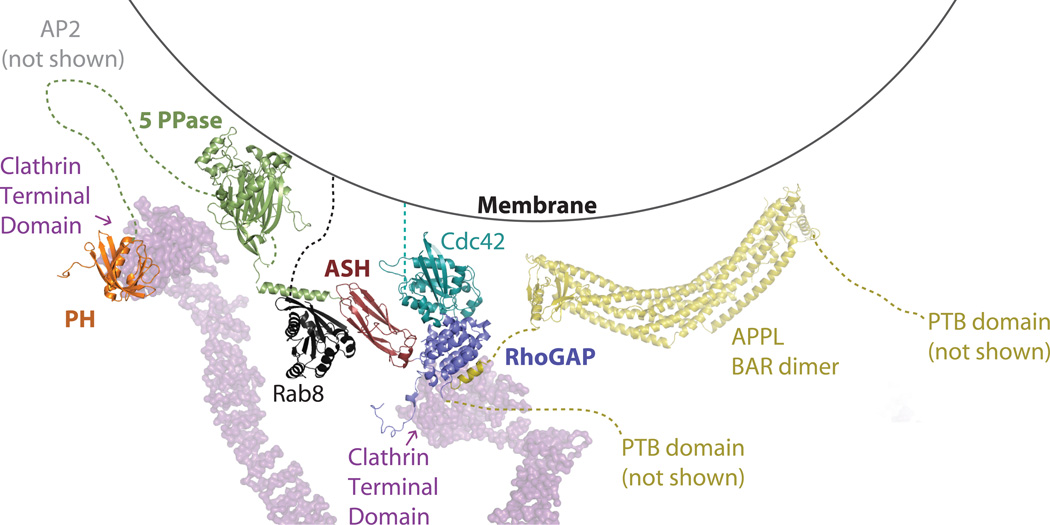
Similar articles
-
OCRL deficiency impairs endolysosomal function in a humanized mouse model for Lowe syndrome and Dent disease.Hum Mol Genet. 2019 Jun 15;28(12):1931-1946. doi: 10.1093/hmg/ddy449. Hum Mol Genet. 2019. PMID: 30590522 Free PMC article.
-
Suppression of intestinal calcium entry channel TRPV6 by OCRL, a lipid phosphatase associated with Lowe syndrome and Dent disease.Am J Physiol Cell Physiol. 2012 May 15;302(10):C1479-91. doi: 10.1152/ajpcell.00277.2011. Epub 2012 Feb 29. Am J Physiol Cell Physiol. 2012. PMID: 22378746 Free PMC article.
-
OCRL-mutated fibroblasts from patients with Dent-2 disease exhibit INPP5B-independent phenotypic variability relatively to Lowe syndrome cells.Hum Mol Genet. 2015 Feb 15;24(4):994-1006. doi: 10.1093/hmg/ddu514. Epub 2014 Oct 9. Hum Mol Genet. 2015. PMID: 25305077
-
The 5-phosphatase OCRL in Lowe syndrome and Dent disease 2.Nat Rev Nephrol. 2017 Aug;13(8):455-470. doi: 10.1038/nrneph.2017.83. Epub 2017 Jul 3. Nat Rev Nephrol. 2017. PMID: 28669993 Review.
-
The cellular and physiological functions of the Lowe syndrome protein OCRL1.Traffic. 2014 May;15(5):471-87. doi: 10.1111/tra.12160. Epub 2014 Mar 7. Traffic. 2014. PMID: 24499450 Free PMC article. Review.
Cited by
-
Expression, Purification, Crystallisation and X-ray Crystallographic Analysis of a Truncated Form of Human Src Homology 2 Containing Inositol 5-Phosphatase 2.Protein J. 2016 Jun;35(3):225-30. doi: 10.1007/s10930-016-9665-y. Protein J. 2016. PMID: 27170292
-
Dent Disease in Chinese Children and Findings from Heterozygous Mothers: Phenotypic Heterogeneity, Fetal Growth, and 10 Novel Mutations.J Pediatr. 2016 Jul;174:204-210.e1. doi: 10.1016/j.jpeds.2016.04.007. Epub 2016 May 9. J Pediatr. 2016. PMID: 27174143 Free PMC article.
-
Phosphatidylinositol 4-kinases and PI4P metabolism in the nervous system: roles in psychiatric and neurological diseases.Mol Neurobiol. 2013 Feb;47(1):361-72. doi: 10.1007/s12035-012-8358-6. Epub 2012 Oct 10. Mol Neurobiol. 2013. PMID: 23054682 Review.
-
Endocrine and behavioural features of Lowe syndrome and their potential molecular mechanisms.J Med Genet. 2022 Dec;59(12):1171-1178. doi: 10.1136/jmedgenet-2022-108490. Epub 2022 Jul 8. J Med Genet. 2022. PMID: 35803701 Free PMC article.
-
Parkinson Sac Domain Mutation in Synaptojanin 1 Impairs Clathrin Uncoating at Synapses and Triggers Dystrophic Changes in Dopaminergic Axons.Neuron. 2017 Feb 22;93(4):882-896.e5. doi: 10.1016/j.neuron.2017.01.019. Neuron. 2017. PMID: 28231468 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
