

Biogenesis of iron-sulfur clusters in mammalian cells: new insights and relevance to human disease
- PMID: 22382365
- PMCID: PMC3291637
- DOI: 10.1242/dmm.009019
Biogenesis of iron-sulfur clusters in mammalian cells: new insights and relevance to human disease
Abstract
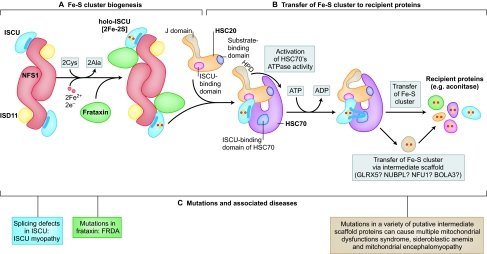
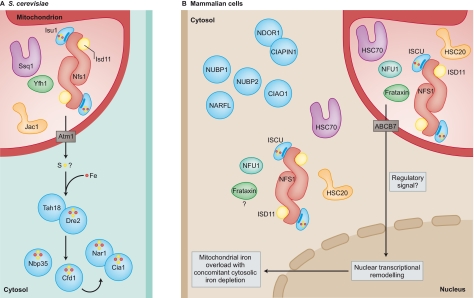
Iron-sulfur (Fe-S) clusters are ubiquitous cofactors composed of iron and inorganic sulfur. They are required for the function of proteins involved in a wide range of activities, including electron transport in respiratory chain complexes, regulatory sensing, photosynthesis and DNA repair. The proteins involved in the biogenesis of Fe-S clusters are evolutionarily conserved from bacteria to humans, and many insights into the process of Fe-S cluster biogenesis have come from studies of model organisms, including bacteria, fungi and plants. It is now clear that several rare and seemingly dissimilar human diseases are attributable to defects in the basic process of Fe-S cluster biogenesis. Although these diseases -which include Friedreich's ataxia (FRDA), ISCU myopathy, a rare form of sideroblastic anemia, an encephalomyopathy caused by dysfunction of respiratory chain complex I and multiple mitochondrial dysfunctions syndrome - affect different tissues, a feature common to many of them is that mitochondrial iron overload develops as a secondary consequence of a defect in Fe-S cluster biogenesis. This Commentary outlines the basic steps of Fe-S cluster biogenesis as they have been defined in model organisms. In addition, it draws attention to refinements of the process that might be specific to the subcellular compartmentalization of Fe-S cluster biogenesis proteins in some eukaryotes, including mammals. Finally, it outlines several important unresolved questions in the field that, once addressed, should offer important clues into how mitochondrial iron homeostasis is regulated, and how dysfunction in Fe-S cluster biogenesis can contribute to disease.
Figures
References
-
- Allikmets R., Raskind W. H., Hutchinson A., Schueck N. D., Dean M., Koeller D. M. (1999). Mutation of a putative mitochondrial iron transporter gene (ABC7) in X-linked sideroblastic anemia and ataxia (XLSA/A). Hum. Mol. Genet. 8, 743–749 - PubMed
-
- Babcock M., de Silva D., Oaks R., Davis-Kaplan S., Jiralerspong S., Montermini L., Pandolfo M., Kaplan J. (1997). Regulation of mitochondrial iron accumulation by Yfh1p, a putative homolog of frataxin. Science 276, 1709–1712 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
