

Engineering naturally occurring trans-acting non-coding RNAs to sense molecular signals
- PMID: 22383579
- PMCID: PMC3384300
- DOI: 10.1093/nar/gks168
Engineering naturally occurring trans-acting non-coding RNAs to sense molecular signals
Abstract
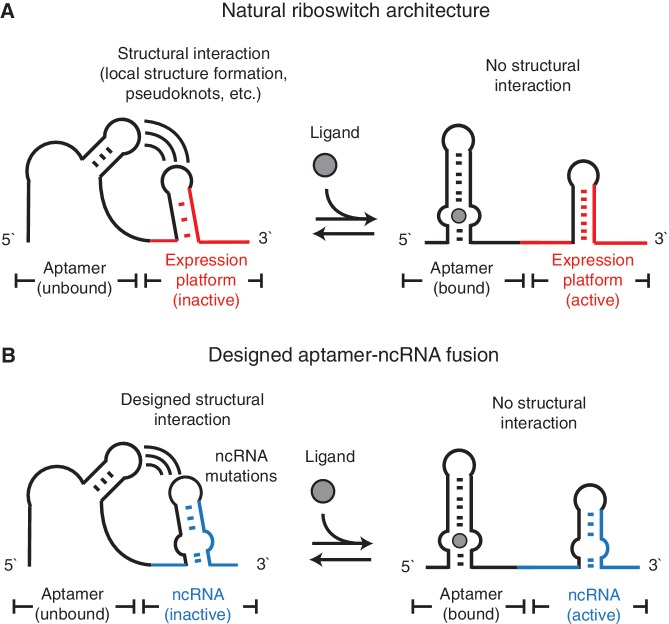
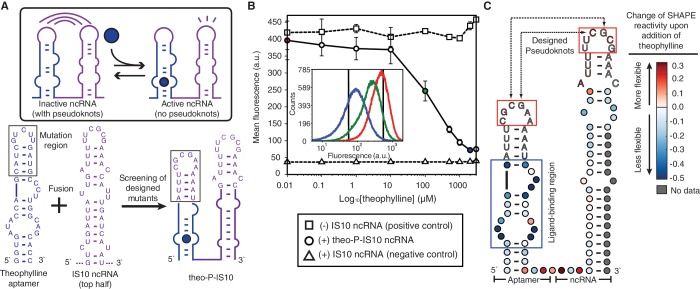
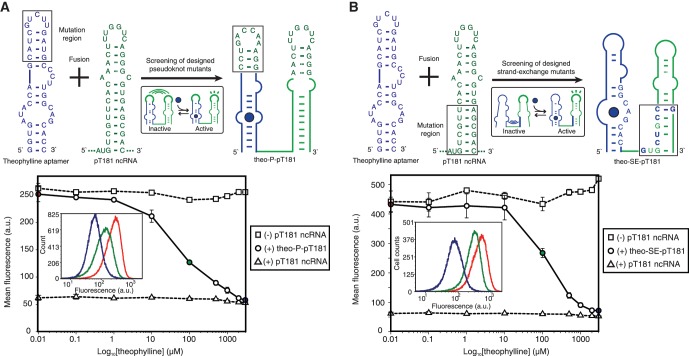
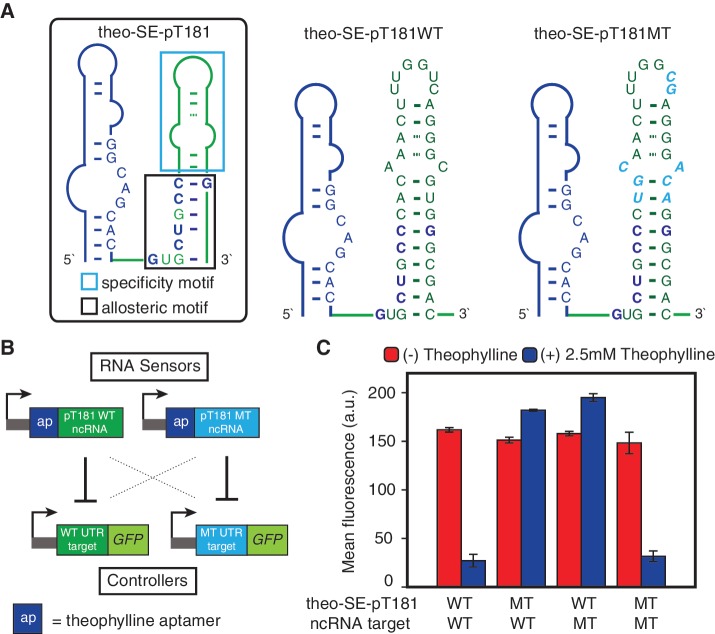
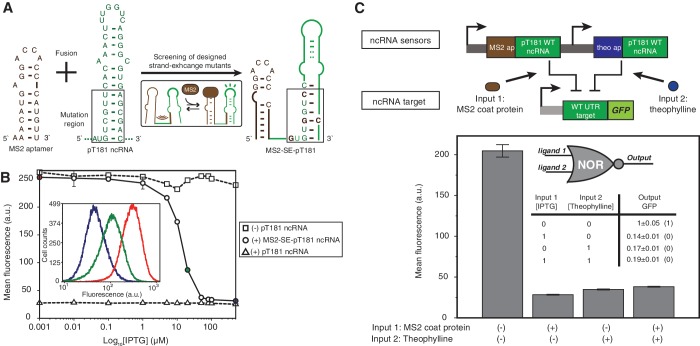
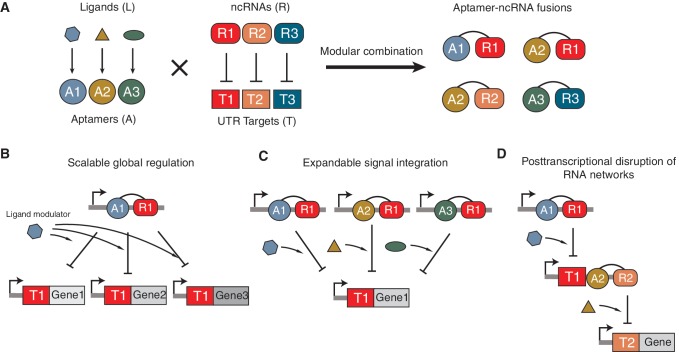
Non-coding RNAs (ncRNAs) are versatile regulators in cellular networks. While most trans-acting ncRNAs possess well-defined mechanisms that can regulate transcription or translation, they generally lack the ability to directly sense cellular signals. In this work, we describe a set of design principles for fusing ncRNAs to RNA aptamers to engineer allosteric RNA fusion molecules that modulate the activity of ncRNAs in a ligand-inducible way in Escherichia coli. We apply these principles to ncRNA regulators that can regulate translation (IS10 ncRNA) and transcription (pT181 ncRNA), and demonstrate that our design strategy exhibits high modularity between the aptamer ligand-sensing motif and the ncRNA target-recognition motif, which allows us to reconfigure these two motifs to engineer orthogonally acting fusion molecules that respond to different ligands and regulate different targets in the same cell. Finally, we show that the same ncRNA fused with different sensing domains results in a sensory-level NOR gate that integrates multiple input signals to perform genetic logic. These ligand-sensing ncRNA regulators provide useful tools to modulate the activity of structurally related families of ncRNAs, and building upon the growing body of RNA synthetic biology, our ability to design aptamer-ncRNA fusion molecules offers new ways to engineer ligand-sensing regulatory circuits.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
