

Chromatin occupancy analysis reveals genome-wide GATA factor switching during hematopoiesis
- PMID: 22383799
- PMCID: PMC3335379
- DOI: 10.1182/blood-2011-09-380634
Chromatin occupancy analysis reveals genome-wide GATA factor switching during hematopoiesis
Abstract
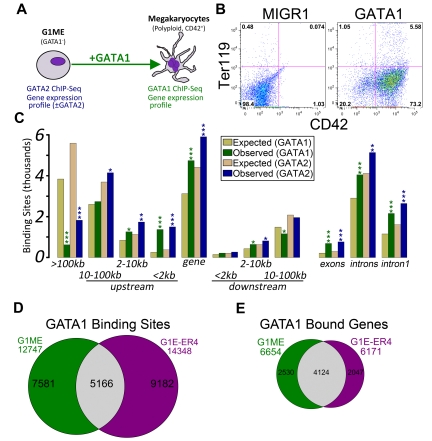
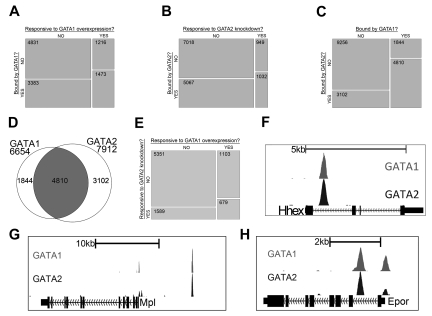
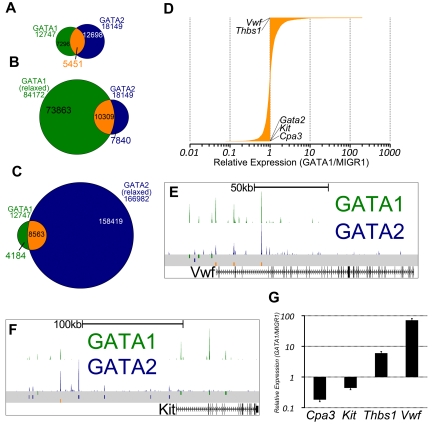
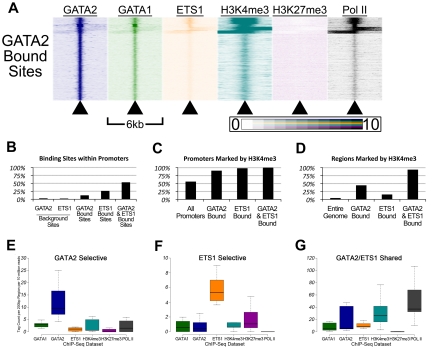
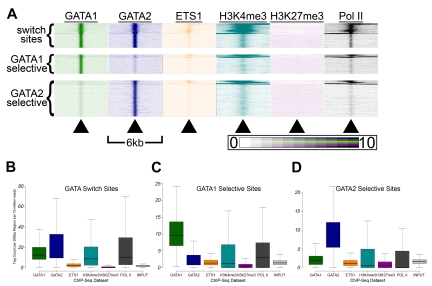
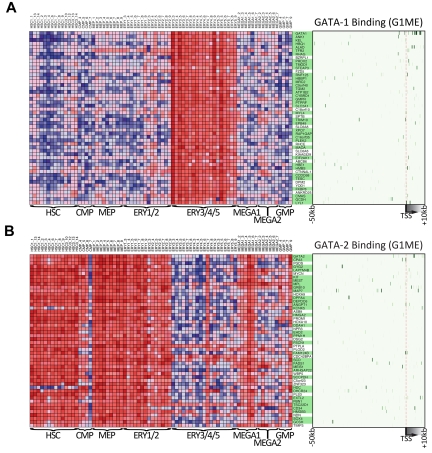
There are many examples of transcription factor families whose members control gene expression profiles of diverse cell types. However, the mechanism by which closely related factors occupy distinct regulatory elements and impart lineage specificity is largely undefined. Here we demonstrate on a genome wide scale that the hematopoietic GATA factors GATA-1 and GATA-2 bind overlapping sets of genes, often at distinct sites, as a means to differentially regulate target gene expression and to regulate the balance between proliferation and differentiation. We also reveal that the GATA switch, which entails a chromatin occupancy exchange between GATA2 and GATA1 in the course of differentiation, operates on more than one-third of GATA1 bound genes. The switch is equally likely to lead to transcriptional activation or repression; and in general, GATA1 and GATA2 act oppositely on switch target genes. In addition, we show that genomic regions co-occupied by GATA2 and the ETS factor ETS1 are strongly enriched for regions marked by H3K4me3 and occupied by Pol II. Finally, by comparing GATA1 occupancy in erythroid cells and megakaryocytes, we find that the presence of ETS factor motifs is a major discriminator of megakaryocyte versus red cell specification.
Figures

Similar articles
-
GATA factor switching from GATA2 to GATA1 contributes to erythroid differentiation.Genes Cells. 2013 Nov;18(11):921-33. doi: 10.1111/gtc.12086. Epub 2013 Aug 1. Genes Cells. 2013. PMID: 23911012
-
GATA-1-mediated transcriptional repression yields persistent transcription factor IIB-chromatin complexes.J Biol Chem. 2006 Dec 8;281(49):37345-52. doi: 10.1074/jbc.M605774200. Epub 2006 Sep 8. J Biol Chem. 2006. PMID: 16963445
-
Regulation of GATA factor expression is distinct between erythroid and mast cell lineages.Mol Cell Biol. 2012 Dec;32(23):4742-55. doi: 10.1128/MCB.00718-12. Epub 2012 Sep 17. Mol Cell Biol. 2012. PMID: 22988301 Free PMC article.
-
A regulatory network governing Gata1 and Gata2 gene transcription orchestrates erythroid lineage differentiation.Int J Hematol. 2014 Nov;100(5):417-24. doi: 10.1007/s12185-014-1568-0. Epub 2014 Mar 18. Int J Hematol. 2014. PMID: 24638828 Review.
-
GATA-related hematologic disorders.Exp Hematol. 2016 Aug;44(8):696-705. doi: 10.1016/j.exphem.2016.05.010. Epub 2016 May 25. Exp Hematol. 2016. PMID: 27235756 Review.
Cited by
-
The Hematopoietic Stem and Progenitor Cell Cistrome: GATA Factor-Dependent cis-Regulatory Mechanisms.Curr Top Dev Biol. 2016;118:45-76. doi: 10.1016/bs.ctdb.2016.01.002. Epub 2016 Feb 26. Curr Top Dev Biol. 2016. PMID: 27137654 Free PMC article. Review.
-
GATA2-induced silencing and LIM-homeodomain protein-induced activation are mediated by a bi-functional response element in the rat GnRH receptor gene.Mol Endocrinol. 2013 Jan;27(1):74-91. doi: 10.1210/me.2012-1182. Epub 2012 Dec 4. Mol Endocrinol. 2013. PMID: 23211524 Free PMC article.
-
A Novel GATA2 Protein Reporter Mouse Reveals Hematopoietic Progenitor Cell Types.Stem Cell Reports. 2020 Aug 11;15(2):326-339. doi: 10.1016/j.stemcr.2020.06.008. Epub 2020 Jul 9. Stem Cell Reports. 2020. PMID: 32649900 Free PMC article.
-
Developmental control of polycomb subunit composition by GATA factors mediates a switch to non-canonical functions.Mol Cell. 2015 Jan 22;57(2):304-316. doi: 10.1016/j.molcel.2014.12.009. Epub 2015 Jan 8. Mol Cell. 2015. PMID: 25578878 Free PMC article.
-
[Clinical and molecular characteristics of GATA2 related pediatric primary myelodysplastic syndrome].Zhonghua Xue Ye Xue Za Zhi. 2019 Jun 14;40(6):477-483. doi: 10.3760/cma.j.issn.0253-2727.2019.06.006. Zhonghua Xue Ye Xue Za Zhi. 2019. PMID: 31340620 Free PMC article. Chinese.
References
-
- Tsai FY, Keller G, Kuo FC, et al. An early haematopoietic defect in mice lacking the transcription factor GATA-2. Nature. 1994;371(6494):221–226. - PubMed
-
- Tsai FY, Orkin SH. Transcription factor GATA-2 is required for proliferation/survival of early hematopoietic cells and mast cell formation, but not for erythroid and myeloid terminal differentiation. Blood. 1997;89(10):3636–3643. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
