

Interleukin-22 drives endogenous thymic regeneration in mice
- PMID: 22383805
- PMCID: PMC3616391
- DOI: 10.1126/science.1218004
Interleukin-22 drives endogenous thymic regeneration in mice
Abstract
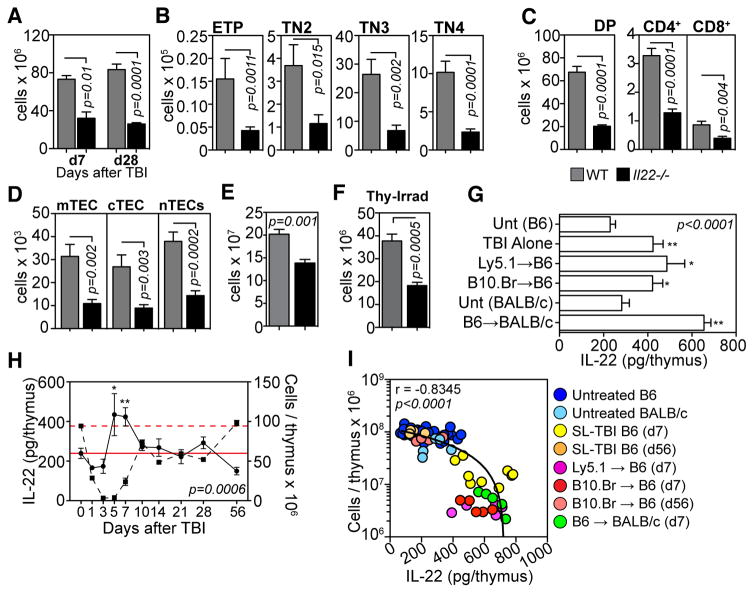
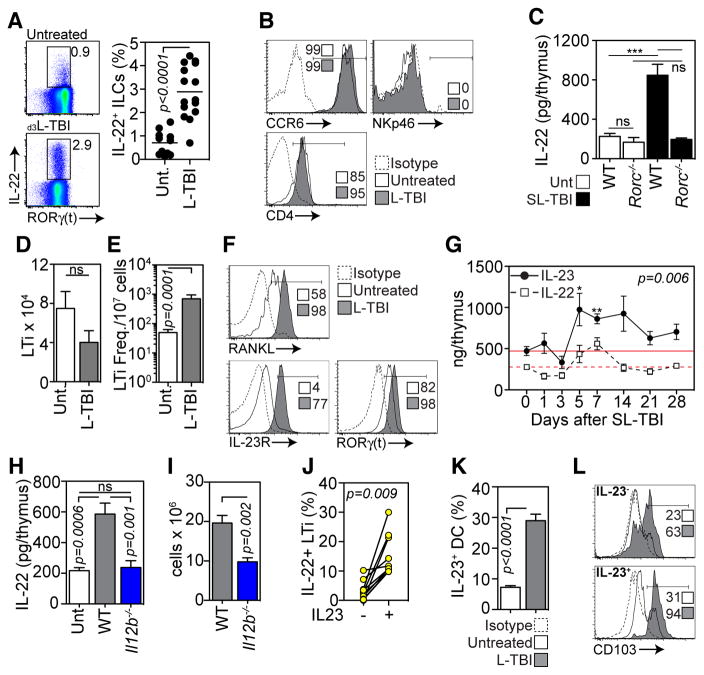
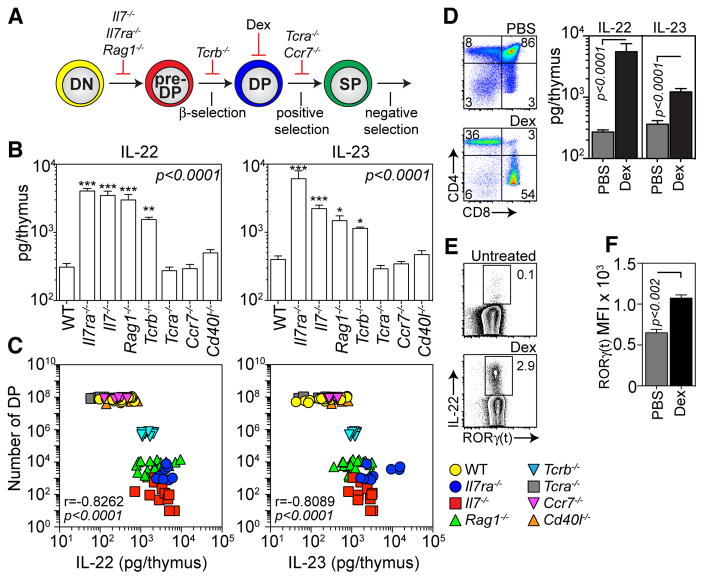
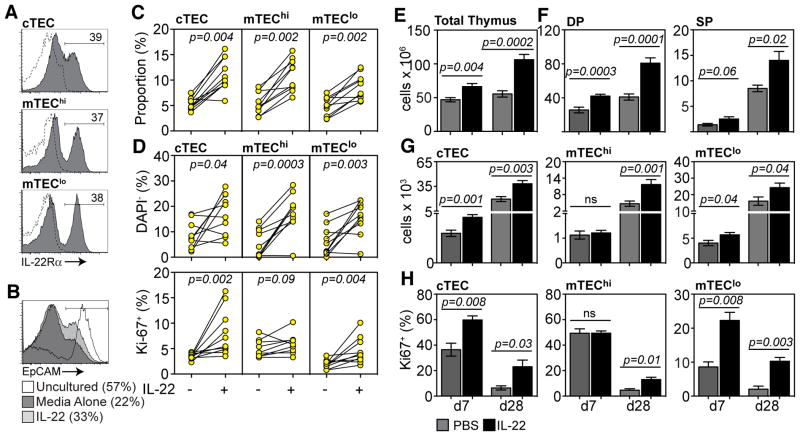
Endogenous thymic regeneration is a crucial function that allows for renewal of immune competence after stress, infection, or immunodepletion. However, the mechanisms governing this regeneration remain poorly understood. We detail such a mechanism, centered on interleukin-22 (IL-22) and triggered by the depletion of CD4(+)CD8(+) double-positive thymocytes. Intrathymic levels of IL-22 were increased after thymic insult, and thymic recovery was impaired in IL-22-deficient mice. IL-22, which signaled through thymic epithelial cells and promoted their proliferation and survival, was up-regulated by radio-resistant RORγ(t)(+)CCR6(+)NKp46(-) lymphoid tissue inducer cells after thymic injury in an IL-23-dependent manner. Administration of IL-22 enhanced thymic recovery after total body irradiation. These studies reveal mechanisms of endogenous thymic repair and offer innovative regenerative strategies for improving immune competence.
Figures
Comment in
-
Immunology. Rebuilding the thymus.Science. 2012 Apr 6;336(6077):40-1. doi: 10.1126/science.1221677. Science. 2012. PMID: 22491841 No abstract available.
-
IL-22 as key factor of thymic regeneration.Immunotherapy. 2012 Jul;4(7):668. Immunotherapy. 2012. PMID: 23019744 No abstract available.
References
-
- van den Brink MR, Alpdogan O, Boyd RL. Strategies to enhance T-cell reconstitution in immunocompromised patients. Nat Rev Immunol. 2004 Nov;4:856. - PubMed
-
- Storek J, Witherspoon RP, Storb R. T cell reconstitution after bone marrow transplantation into adult patients does not resemble T cell development in early life. Bone Marrow Transplant. 1995 Sep;16:413. - PubMed
-
- Takahama Y. Journey through the thymus: stromal guides for T-cell development and selection. Nat Rev Immunol. 2006 Feb;6:127. - PubMed
-
- Petrie HT, Zuniga-Pflucker JC. Zoned out: functional mapping of stromal signaling microenvironments in the thymus. Annu Rev Immunol. 2007;25:649. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
