

A pathogenic mechanism in Huntington's disease involves small CAG-repeated RNAs with neurotoxic activity
- PMID: 22383888
- PMCID: PMC3285580
- DOI: 10.1371/journal.pgen.1002481
A pathogenic mechanism in Huntington's disease involves small CAG-repeated RNAs with neurotoxic activity
Abstract
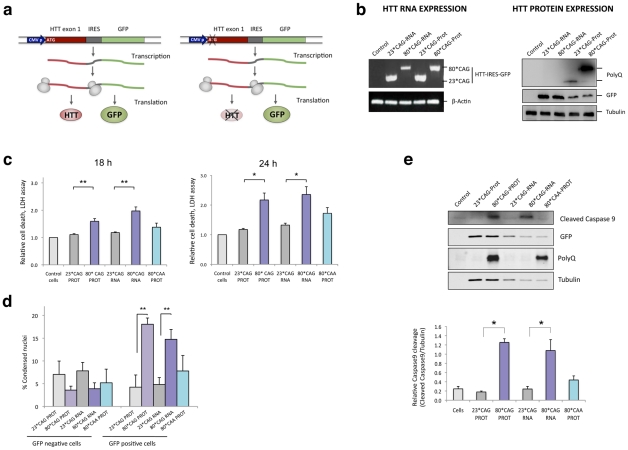
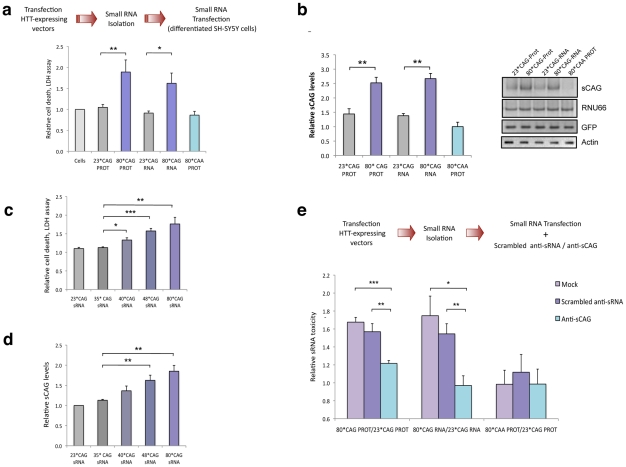
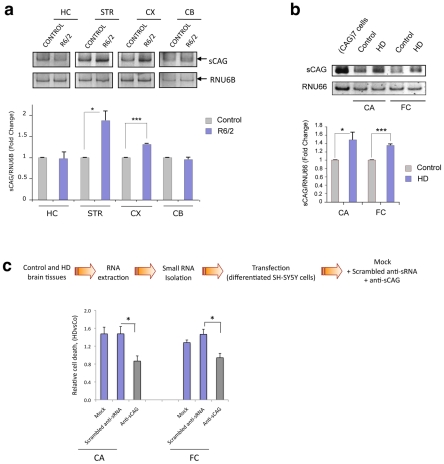
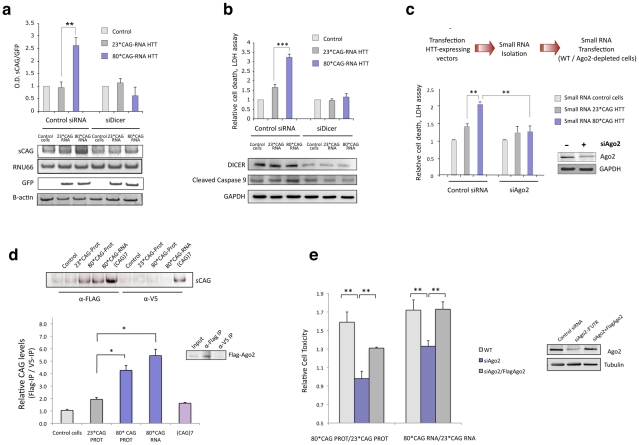
Huntington's disease (HD) is an autosomal dominantly inherited disorder caused by the expansion of CAG repeats in the Huntingtin (HTT) gene. The abnormally extended polyglutamine in the HTT protein encoded by the CAG repeats has toxic effects. Here, we provide evidence to support that the mutant HTT CAG repeats interfere with cell viability at the RNA level. In human neuronal cells, expanded HTT exon-1 mRNA with CAG repeat lengths above the threshold for complete penetrance (40 or greater) induced cell death and increased levels of small CAG-repeated RNAs (sCAGs), of ≈21 nucleotides in a Dicer-dependent manner. The severity of the toxic effect of HTT mRNA and sCAG generation correlated with CAG expansion length. Small RNAs obtained from cells expressing mutant HTT and from HD human brains significantly decreased neuronal viability, in an Ago2-dependent mechanism. In both cases, the use of anti-miRs specific for sCAGs efficiently blocked the toxic effect, supporting a key role of sCAGs in HTT-mediated toxicity. Luciferase-reporter assays showed that expanded HTT silences the expression of CTG-containing genes that are down-regulated in HD. These results suggest a possible link between HD and sCAG expression with an aberrant activation of the siRNA/miRNA gene silencing machinery, which may trigger a detrimental response. The identification of the specific cellular processes affected by sCAGs may provide insights into the pathogenic mechanisms underlying HD, offering opportunities to develop new therapeutic approaches.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
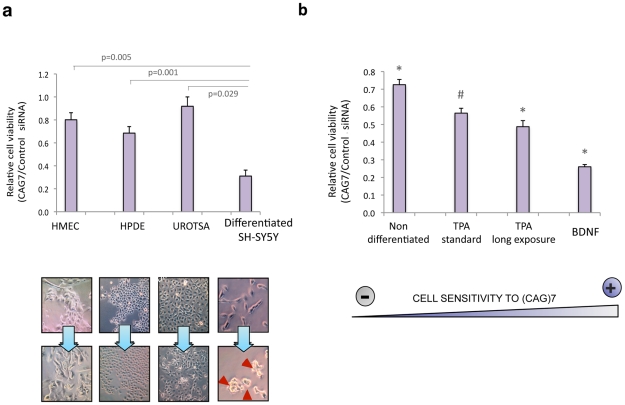
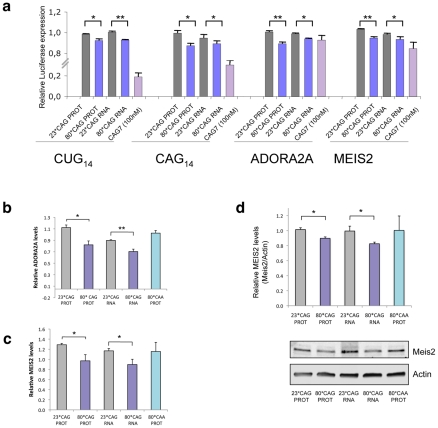
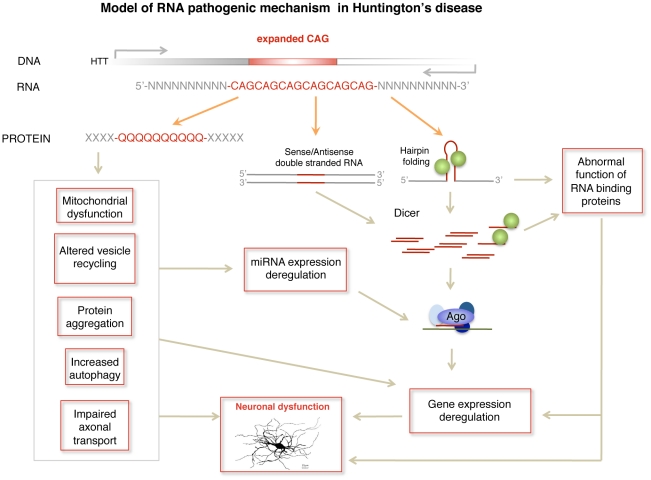
Comment in
-
Diced triplets expose neurons to RISC.PLoS Genet. 2012;8(2):e1002545. doi: 10.1371/journal.pgen.1002545. Epub 2012 Feb 23. PLoS Genet. 2012. PMID: 22383898 Free PMC article. No abstract available.
Similar articles
-
RNA toxicity induced by expanded CAG repeats in Huntington's disease.Brain Pathol. 2016 Nov;26(6):779-786. doi: 10.1111/bpa.12427. Brain Pathol. 2016. PMID: 27529325 Free PMC article. Review.
-
The length of uninterrupted CAG repeats in stem regions of repeat disease associated hairpins determines the amount of short CAG oligonucleotides that are toxic to cells through RNA interference.Cell Death Dis. 2022 Dec 30;13(12):1078. doi: 10.1038/s41419-022-05494-1. Cell Death Dis. 2022. PMID: 36585400 Free PMC article.
-
Phosphorodiamidate morpholino oligomers suppress mutant huntingtin expression and attenuate neurotoxicity.Hum Mol Genet. 2014 Dec 1;23(23):6302-17. doi: 10.1093/hmg/ddu349. Epub 2014 Jul 4. Hum Mol Genet. 2014. PMID: 25035419 Free PMC article.
-
Targeting CAG repeat RNAs reduces Huntington's disease phenotype independently of huntingtin levels.J Clin Invest. 2016 Nov 1;126(11):4319-4330. doi: 10.1172/JCI83185. Epub 2016 Oct 10. J Clin Invest. 2016. PMID: 27721240 Free PMC article.
-
Multiple clinical features of Huntington's disease correlate with mutant HTT gene CAG repeat lengths and neurodegeneration.J Neurol. 2019 Mar;266(3):551-564. doi: 10.1007/s00415-018-8940-6. Epub 2018 Jun 28. J Neurol. 2019. PMID: 29956026 Review.
Cited by
-
Nuclear retention of full-length HTT RNA is mediated by splicing factors MBNL1 and U2AF65.Sci Rep. 2015 Jul 28;5:12521. doi: 10.1038/srep12521. Sci Rep. 2015. PMID: 26218986 Free PMC article.
-
RNA-mediated pathogenic mechanisms in polyglutamine diseases and amyotrophic lateral sclerosis.Front Cell Neurosci. 2014 Dec 19;8:431. doi: 10.3389/fncel.2014.00431. eCollection 2014. Front Cell Neurosci. 2014. PMID: 25565965 Free PMC article. Review.
-
CAG RNAs induce DNA damage and apoptosis by silencing NUDT16 expression in polyglutamine degeneration.Proc Natl Acad Sci U S A. 2021 May 11;118(19):e2022940118. doi: 10.1073/pnas.2022940118. Proc Natl Acad Sci U S A. 2021. PMID: 33947817 Free PMC article.
-
Oligonucleotides Targeting DNA Repeats Downregulate Huntingtin Gene Expression in Huntington's Patient-Derived Neural Model System.Nucleic Acid Ther. 2021 Dec;31(6):443-456. doi: 10.1089/nat.2021.0021. Epub 2021 Sep 13. Nucleic Acid Ther. 2021. PMID: 34520257 Free PMC article.
-
RAN Translation in Huntington Disease.Neuron. 2015 Nov 18;88(4):667-77. doi: 10.1016/j.neuron.2015.10.038. Neuron. 2015. PMID: 26590344 Free PMC article.
References
-
- Schilling G, Sharp AH, Loev SJ, Wagster MV, Li SH, et al. Expression of the Huntington's disease (IT15) protein product in HD patients. Hum Mol Genet. 1995;4:1365–1371. - PubMed
-
- Sharp AH, Loev SJ, Schilling G, Li SH, Li XJ, et al. Widespread expression of Huntington's disease gene (IT15) protein product. Neuron. 1995;14:1065–1074. - PubMed
-
- Vonsattel JP, DiFiglia M. Huntington disease. J Neuropathol Exp Neurol. 1998;57:369–384. - PubMed
-
- Walker FO. Huntington's disease. Lancet. 2007;369:218–228. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
