

Network deficiency exacerbates impairment in a mouse model of retinal degeneration
- PMID: 22383900
- PMCID: PMC3285818
- DOI: 10.3389/fnsys.2012.00008
Network deficiency exacerbates impairment in a mouse model of retinal degeneration
Abstract
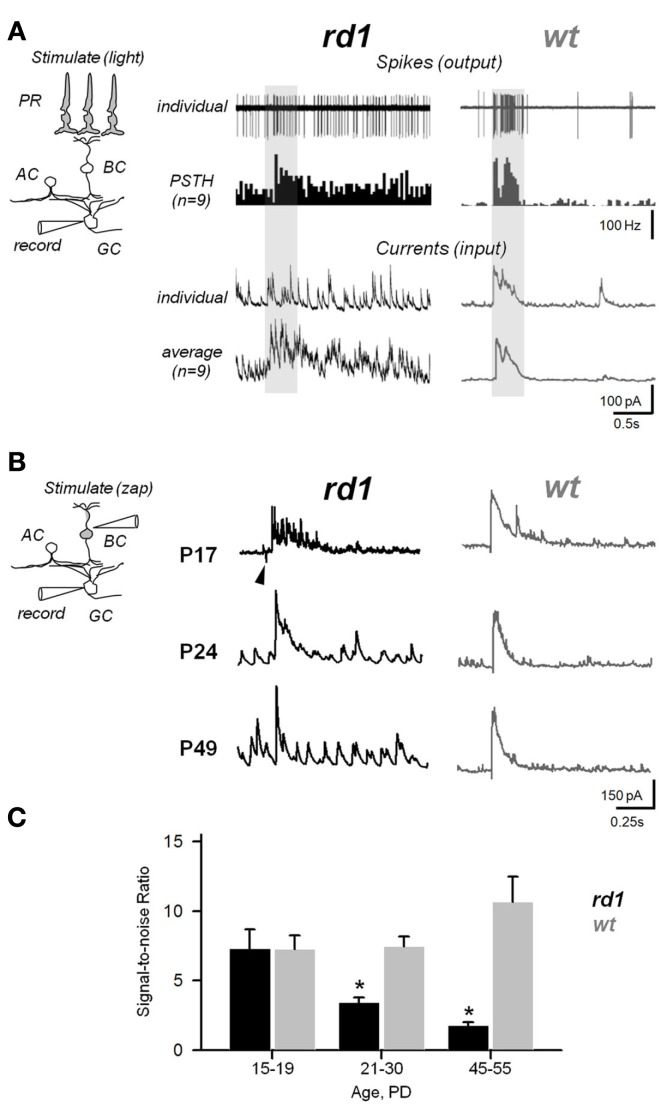
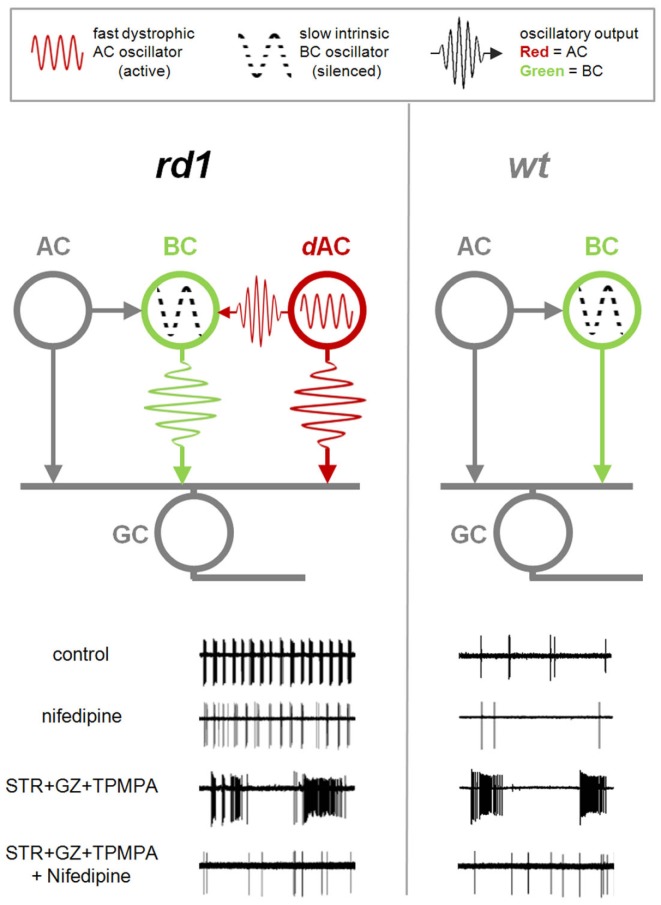
Neural oscillations play an important role in normal brain activity, but also manifest during Parkinson's disease, epilepsy, and other pathological conditions. The contribution of these aberrant oscillations to the function of the surviving brain remains unclear. In recording from retina in a mouse model of retinal degeneration (RD), we found that the incidence of oscillatory activity varied across different cell classes, evidence that some retinal networks are more affected by functional changes than others. This aberrant activity was driven by an independent inhibitory amacrine cell oscillator. By stimulating the surviving circuitry at different stages of the neurodegenerative process, we found that this dystrophic oscillator further compromises the function of the retina. These data reveal that retinal remodeling can exacerbate the visual deficit, and that aberrant synaptic activity could be targeted for RD treatment.
Keywords: functional remodeling; neurodegeneration; oscillations; retinitis pigmentosa; synaptic plasticity.
Figures
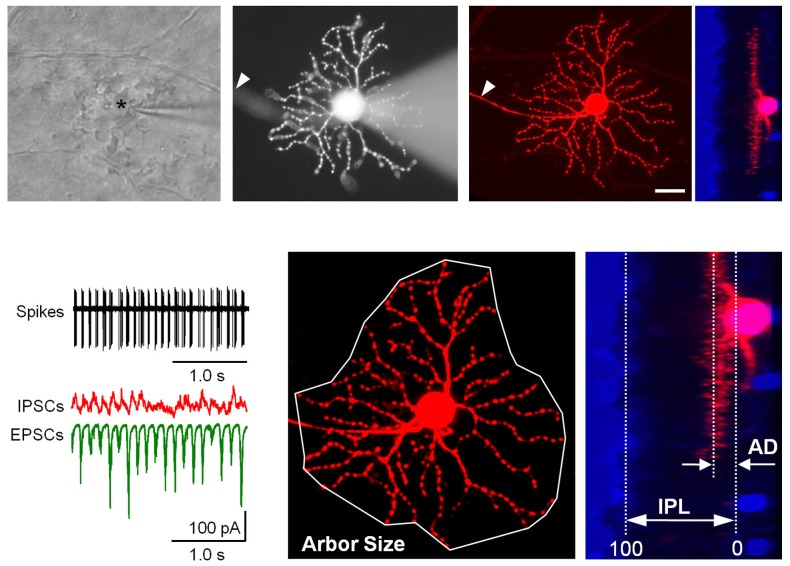
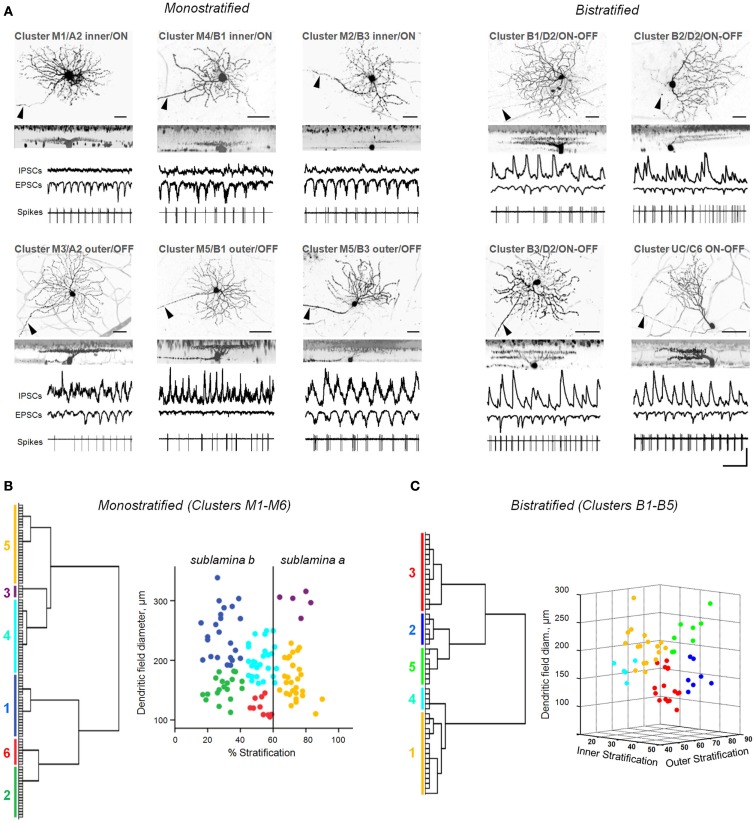
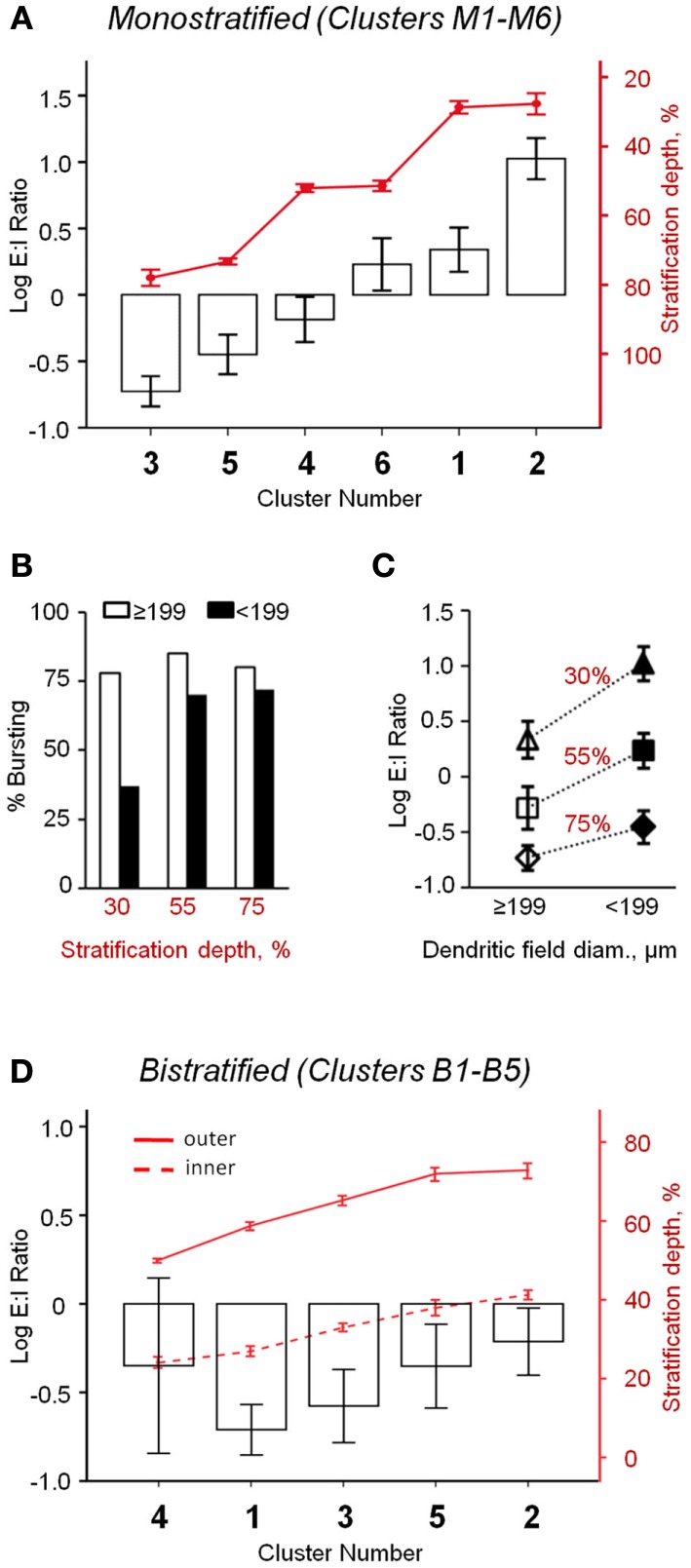
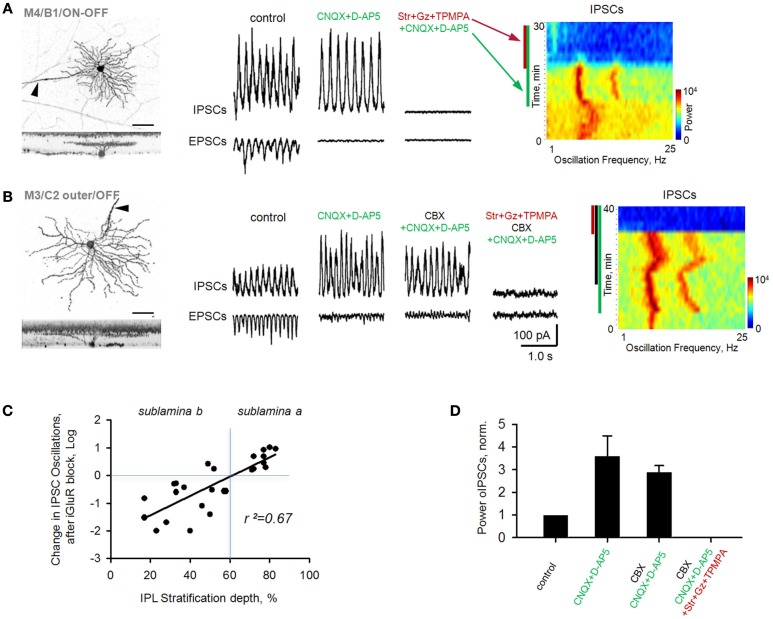
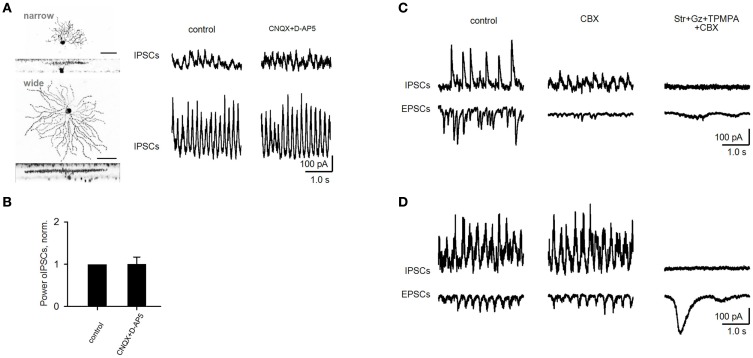
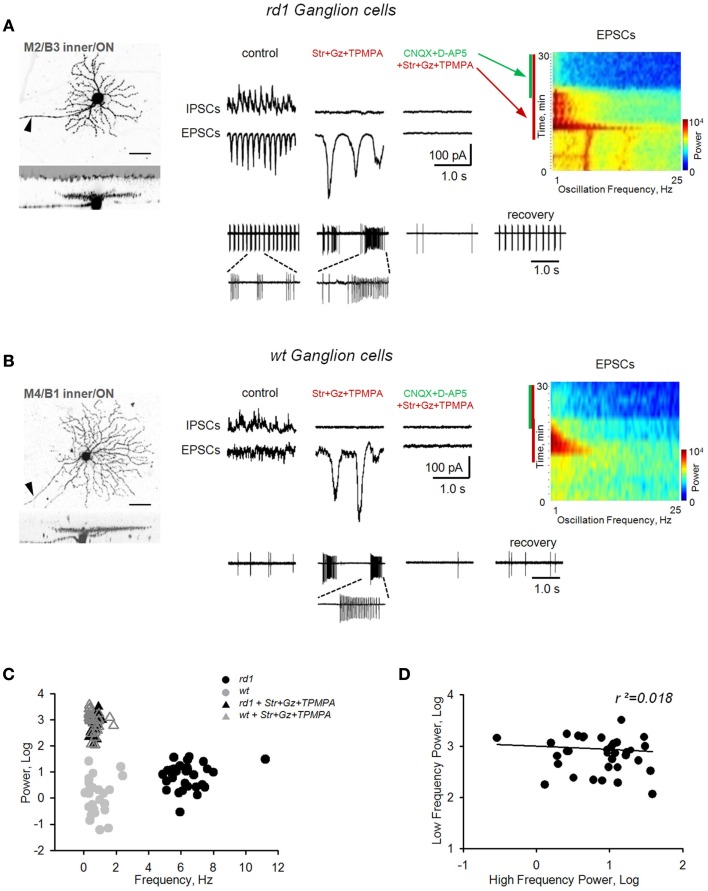
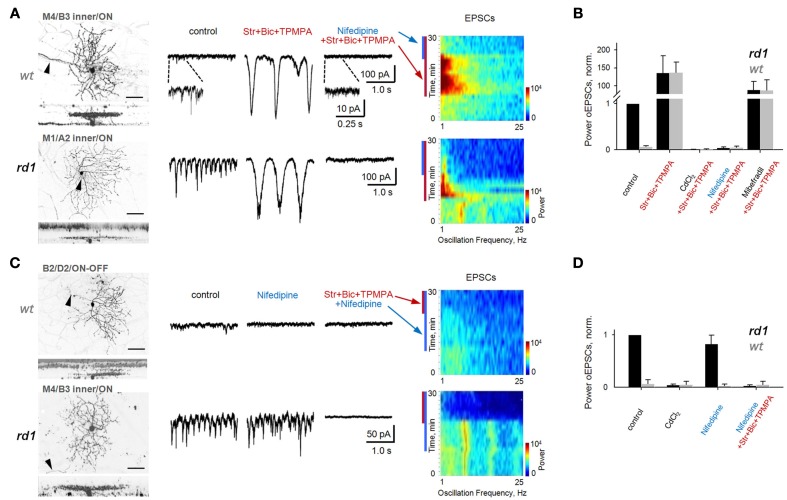
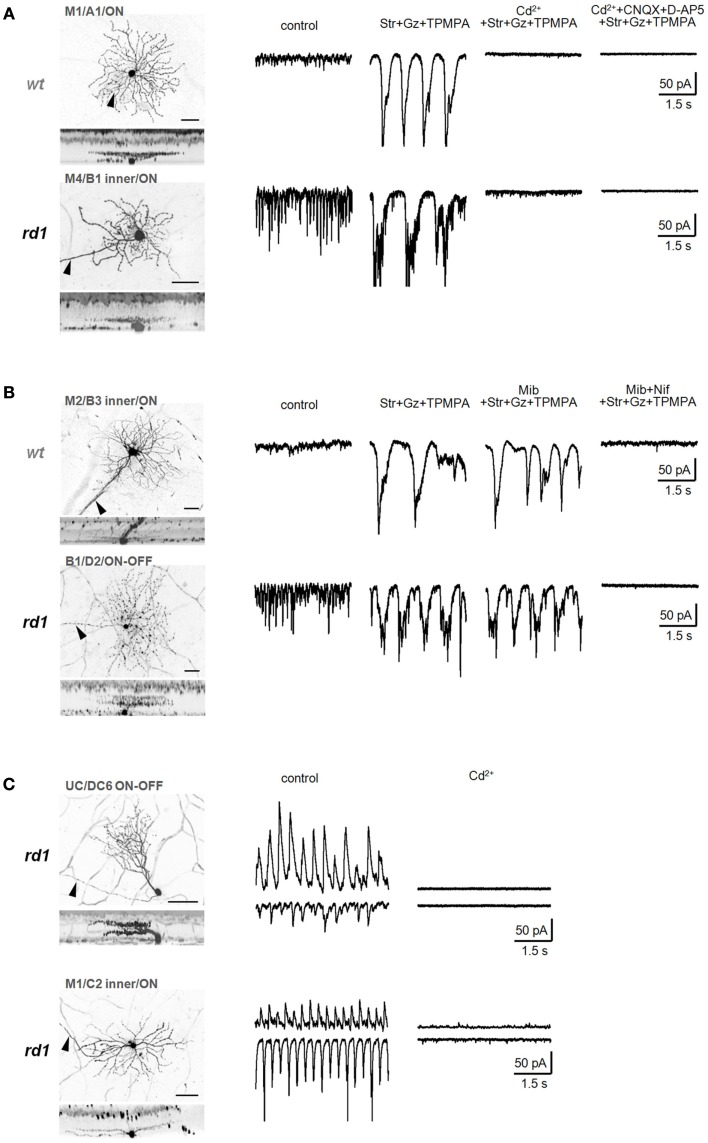
Similar articles
-
Functional remodeling of glutamate receptors by inner retinal neurons occurs from an early stage of retinal degeneration.J Comp Neurol. 2009 Jun 10;514(5):473-91. doi: 10.1002/cne.22029. J Comp Neurol. 2009. PMID: 19350664
-
Disruption in dopaminergic innervation during photoreceptor degeneration.J Comp Neurol. 2016 Apr 15;524(6):1208-21. doi: 10.1002/cne.23899. Epub 2015 Sep 28. J Comp Neurol. 2016. PMID: 26356010 Free PMC article.
-
Activation of Rod Input in a Model of Retinal Degeneration Reverses Retinal Remodeling and Induces Formation of Functional Synapses and Recovery of Visual Signaling in the Adult Retina.J Neurosci. 2019 Aug 21;39(34):6798-6810. doi: 10.1523/JNEUROSCI.2902-18.2019. Epub 2019 Jul 8. J Neurosci. 2019. PMID: 31285302 Free PMC article.
-
Multiple Independent Oscillatory Networks in the Degenerating Retina.Front Cell Neurosci. 2015 Nov 9;9:444. doi: 10.3389/fncel.2015.00444. eCollection 2015. Front Cell Neurosci. 2015. PMID: 26617491 Free PMC article. Review.
-
Degeneration-Dependent Retinal Remodeling: Looking for the Molecular Trigger.Front Neurosci. 2020 Dec 18;14:618019. doi: 10.3389/fnins.2020.618019. eCollection 2020. Front Neurosci. 2020. PMID: 33390897 Free PMC article. Review.
Cited by
-
Aberrant activity in retinal degeneration impairs central visual processing and relies on Cx36-containing gap junctions.Exp Eye Res. 2016 Sep;150:81-9. doi: 10.1016/j.exer.2015.05.013. Epub 2015 May 22. Exp Eye Res. 2016. PMID: 26005040 Free PMC article.
-
Elucidating the role of AII amacrine cells in glutamatergic retinal waves.J Neurosci. 2015 Jan 28;35(4):1675-86. doi: 10.1523/JNEUROSCI.3291-14.2015. J Neurosci. 2015. PMID: 25632142 Free PMC article.
-
Atypical Expression and Activation of GluN2A- and GluN2B-Containing NMDA Receptors at Ganglion Cells during Retinal Degeneration.Neuroscience. 2018 Nov 21;393:61-72. doi: 10.1016/j.neuroscience.2018.09.048. Epub 2018 Oct 10. Neuroscience. 2018. PMID: 30312782 Free PMC article.
-
Differences in the spatial fidelity of evoked and spontaneous signals in the degenerating retina.Front Cell Neurosci. 2022 Nov 7;16:1040090. doi: 10.3389/fncel.2022.1040090. eCollection 2022. Front Cell Neurosci. 2022. PMID: 36419935 Free PMC article.
-
Retinal ganglion cells undergo cell type-specific functional changes in a computational model of cone-mediated retinal degeneration.Front Neurosci. 2023 May 18;17:1147729. doi: 10.3389/fnins.2023.1147729. eCollection 2023. Front Neurosci. 2023. PMID: 37274203 Free PMC article.
References
-
- Borowska J. T., Trenholm S., Awatramani G. B. (2010). Intrinsic mechanisms in bipolar cells drive spontaneous network activity during retinal degeneration. ARVO Meet Abstr. 51, 2486.
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous