

Mosaic origins of a complex chimeric mitochondrial gene in Silene vulgaris
- PMID: 22383961
- PMCID: PMC3288002
- DOI: 10.1371/journal.pone.0030401
Mosaic origins of a complex chimeric mitochondrial gene in Silene vulgaris
Abstract
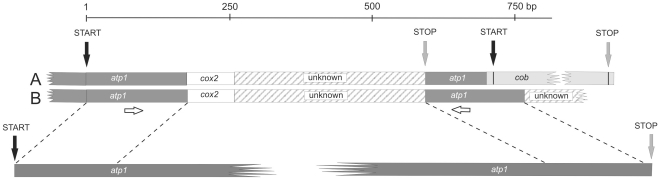
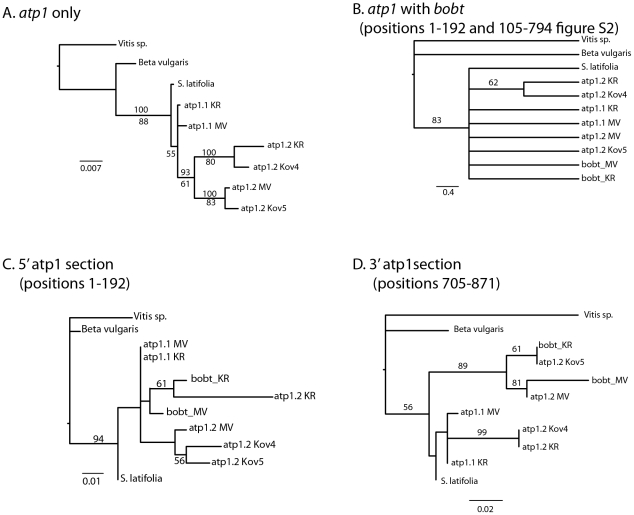
Chimeric genes are significant sources of evolutionary innovation that are normally created when portions of two or more protein coding regions fuse to form a new open reading frame. In plant mitochondria astonishingly high numbers of different novel chimeric genes have been reported, where they are generated through processes of rearrangement and recombination. Nonetheless, because most studies do not find or report nucleotide variation within the same chimeric gene, evolution after the origination of these chimeric genes remains unstudied. Here we identify two alleles of a complex chimera in Silene vulgaris that are divergent in nucleotide sequence, genomic position relative to other mitochondrial genes, and expression patterns. Structural patterns suggest a history partially influenced by gene conversion between the chimeric gene and functional copies of subunit 1 of the mitochondrial ATP synthase gene (atp1). We identified small repeat structures within the chimeras that are likely recombination sites allowing generation of the chimera. These results establish the potential for chimeric gene divergence in different plant mitochondrial lineages within the same species. This result contrasts with the absence of diversity within mitochondrial chimeras found in crop species.
Conflict of interest statement
Figures



Similar articles
-
Transcription of atp1 is influenced by both genomic configuration and nuclear background in the highly rearranged mitochondrial genomes of Silene vulgaris.Plant Mol Biol. 2013 Mar;81(4-5):495-505. doi: 10.1007/s11103-013-0018-3. Epub 2013 Jan 30. Plant Mol Biol. 2013. PMID: 23361622
-
Variation in mutation rate and polymorphism among mitochondrial genes of Silene vulgaris.Mol Biol Evol. 2007 Aug;24(8):1783-91. doi: 10.1093/molbev/msm106. Epub 2007 May 28. Mol Biol Evol. 2007. PMID: 17533174
-
Nonneutral evolution of organelle genes in Silene vulgaris.Genetics. 2006 Dec;174(4):1983-94. doi: 10.1534/genetics.106.060202. Epub 2006 Sep 15. Genetics. 2006. PMID: 16980398 Free PMC article.
-
Origination of chimeric genes through DNA-level recombination.Genome Dyn. 2007;3:131-146. doi: 10.1159/000107608. Genome Dyn. 2007. PMID: 18753789 Review.
-
Chimerism: A whole new perspective in gene regulation.Biochim Biophys Acta Gen Subj. 2025 Mar;1869(3):130767. doi: 10.1016/j.bbagen.2025.130767. Epub 2025 Jan 22. Biochim Biophys Acta Gen Subj. 2025. PMID: 39855315 Review.
Cited by
-
Differentially Expressed Genes Shared by Two Distinct Cytoplasmic Male Sterility (CMS) Types of Silene vulgaris Suggest the Importance of Oxidative Stress in Pollen Abortion.Cells. 2020 Dec 16;9(12):2700. doi: 10.3390/cells9122700. Cells. 2020. PMID: 33339225 Free PMC article.
-
Next-generation sequencing of mitochondrial targeted AAV transfer of human ND4 in mice.Mol Vis. 2013 Jul 14;19:1482-91. Print 2013. Mol Vis. 2013. PMID: 23869167 Free PMC article.
-
Transcription of atp1 is influenced by both genomic configuration and nuclear background in the highly rearranged mitochondrial genomes of Silene vulgaris.Plant Mol Biol. 2013 Mar;81(4-5):495-505. doi: 10.1007/s11103-013-0018-3. Epub 2013 Jan 30. Plant Mol Biol. 2013. PMID: 23361622
-
Homologous recombination changes the context of Cytochrome b transcription in the mitochondrial genome of Silene vulgaris KRA.BMC Genomics. 2018 Dec 4;19(1):874. doi: 10.1186/s12864-018-5254-0. BMC Genomics. 2018. PMID: 30514207 Free PMC article.
-
Non-coding RNA may be associated with cytoplasmic male sterility in Silene vulgaris.J Exp Bot. 2017 Mar 1;68(7):1599-1612. doi: 10.1093/jxb/erx057. J Exp Bot. 2017. PMID: 28369520 Free PMC article.
References
-
- Katju V, Lynch M. On the formation of novel genes by duplication in the caenorhabditis elegans genome. Molecular Biology and Evolution. 2006;23:1056–1067. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
