

Cognitive control reflects context monitoring, not motoric stopping, in response inhibition
- PMID: 22384038
- PMCID: PMC3288048
- DOI: 10.1371/journal.pone.0031546
Cognitive control reflects context monitoring, not motoric stopping, in response inhibition
Abstract
The inhibition of unwanted behaviors is considered an effortful and controlled ability. However, inhibition also requires the detection of contexts indicating that old behaviors may be inappropriate--in other words, inhibition requires the ability to monitor context in the service of goals, which we refer to as context-monitoring. Using behavioral, neuroimaging, electrophysiological and computational approaches, we tested whether motoric stopping per se is the cognitively-controlled process supporting response inhibition, or whether context-monitoring may fill this role. Our results demonstrate that inhibition does not require control mechanisms beyond those involved in context-monitoring, and that such control mechanisms are the same regardless of stopping demands. These results challenge dominant accounts of inhibitory control, which posit that motoric stopping is the cognitively-controlled process of response inhibition, and clarify emerging debates on the frontal substrates of response inhibition by replacing the centrality of controlled mechanisms for motoric stopping with context-monitoring.
Conflict of interest statement
Figures
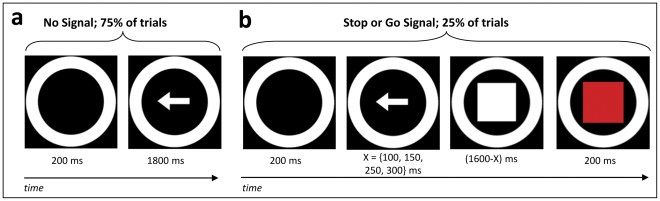
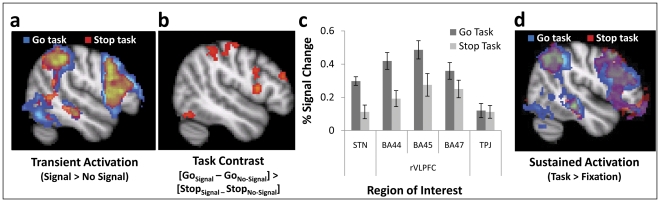
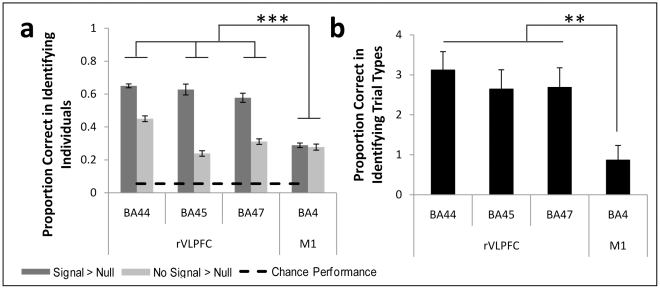
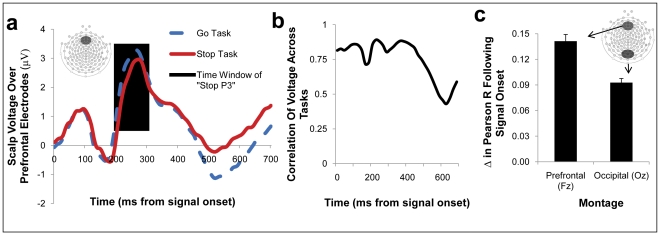
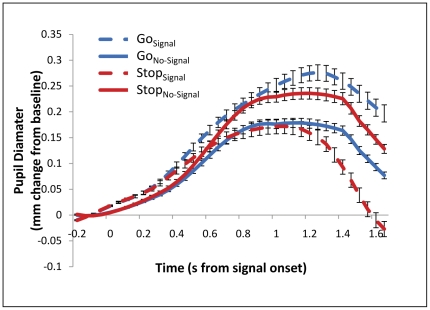
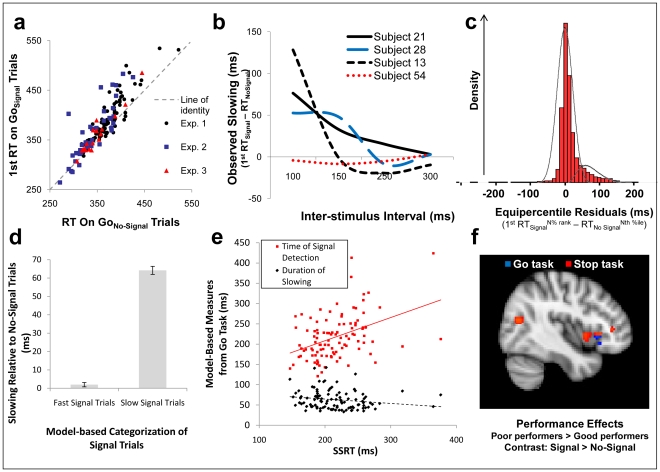
References
-
- Aron AR. The neural basis of inhibition in cognitive control. Neuroscientist. 2007;13:214–28. - PubMed
-
- Aron AR, Poldrack RA. The cognitive neuroscience of response inhibition: Relevance for genetic research in ADHD. Biological Psychiatry, 2005;57:1285–92. - PubMed
-
- MacLeod CM, Dodd MD, Sheard ED, Wilson DE, Bibi U. In Opposition to Inhibition. In: Ross BH, editor. The Psychology of Learning and Motivation. San Diego: Academic Press; 2003. pp. 163–214.
-
- Ferrier D. The functions of the brain. London: Elder; 1876. 353
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
