

Genetic Diversity and Genomic Plasticity of Cryptococcus neoformans AD Hybrid Strains
- PMID: 22384385
- PMCID: PMC3276195
- DOI: 10.1534/g3.111.001255
Genetic Diversity and Genomic Plasticity of Cryptococcus neoformans AD Hybrid Strains
Abstract
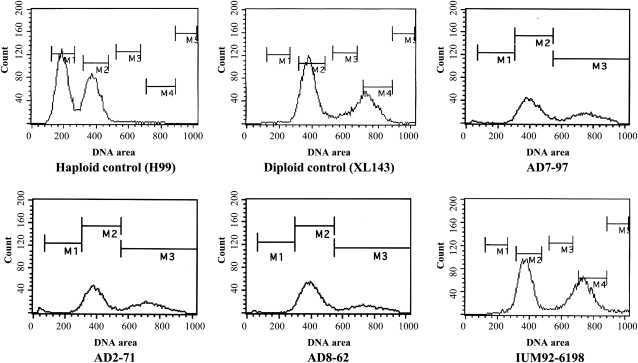
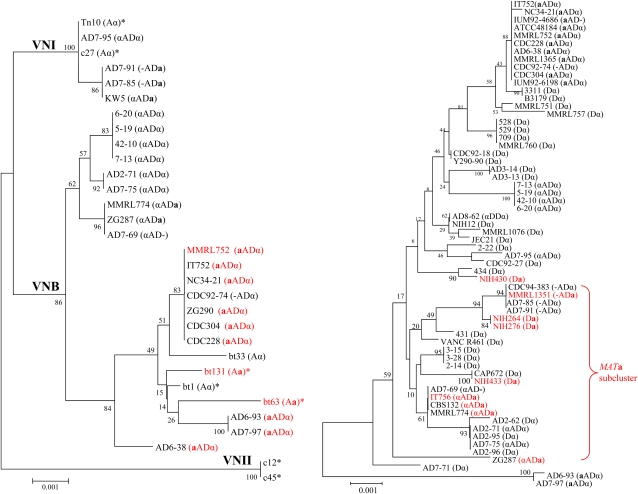
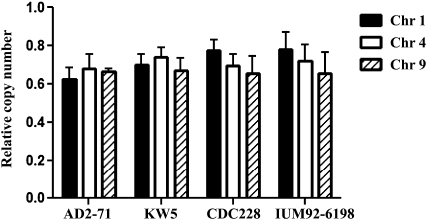
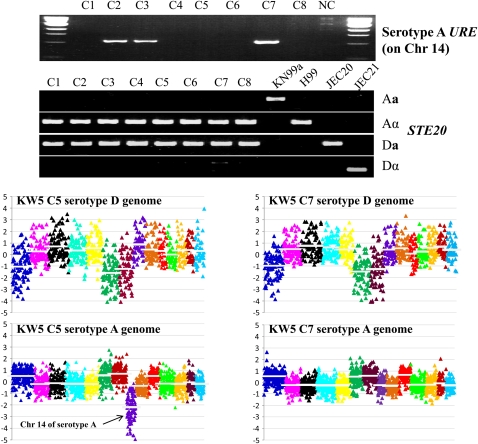
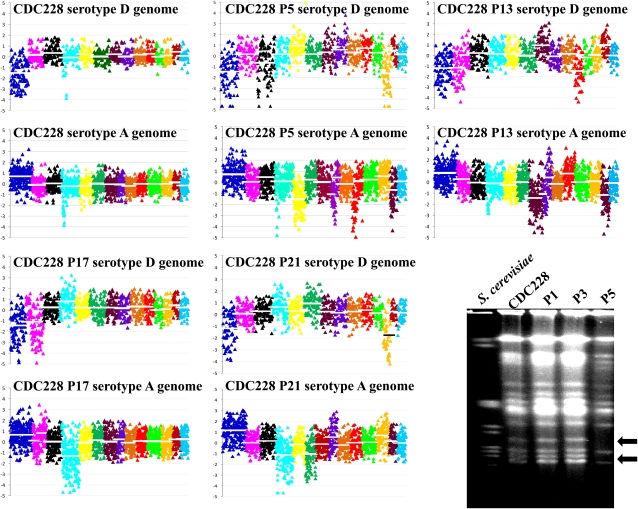
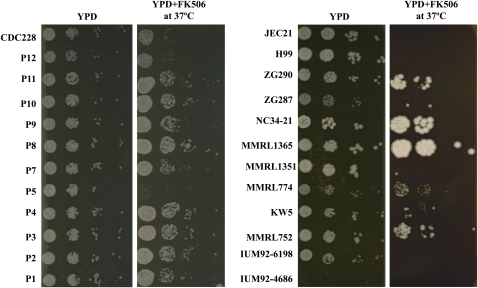
Natural hybridization between two strains, varieties, or species is a common phenomenon in both plants and animals. Although hybridization may skew established gene pools, it generates population diversity efficiently and sometimes results in the emergence of newly adapted genotypes. Cryptococcus neoformans, which causes the most frequent opportunistic fungal infection in immunocompromised hosts, has three serotypes: A, D, and AD. Serotype-specific multilocus sequence typing and serotype-specific comparative genome hybridization were applied to investigate the genetic variability and genomic organization of C. neoformans serotype AD isolates. We confirm that C. neoformans serotype AD isolates are hybrids of serotype A and D strains. Compared with haploid strains, most AD hybrid isolates exhibit unique multilocus sequence typing genotypes, suggesting that multiple independent hybridization events punctuated the origin and evolutionary trajectory of AD hybrids. The MATa alleles from both haploid and AD hybrid isolates group closely to form a cluster or subcluster in both the serotype A and D populations. The rare and unique distribution of MATa alleles may restrict sexual reproduction between isolates of opposite mating types. The genetic diversity of the serotype D population, including haploid strains and serotype D genomes of the AD hybrid, is significantly greater than that of serotype A, and there are signatures of recombination within the serotype D population. Given that MATa isolates are relatively rare, both opposite-sex and same-sex mating may contribute to genetic recombination of serotype D in nature. Extensive chromosome loss was observed in AD hybrid isolates, which results in loss of heterozygosity in the otherwise-heterozygous AD hybrid genome. Most AD hybrid isolates exhibit hybrid vigor and are resistant to the antifungal drug FK506. In addition, the C. neoformans AD hybrid genome is highly dynamic, with continuous chromosome loss, which may be a facile route for pathogen evolution through which genotypic and phenotypic variation is generated.
Figures

References
-
- Abbott R.A., Lowe A. J., 2004. Origins, establishment and evolution of new polyploid species: Senecio cambrensis and S. eboracensis in the British Isles. Biol. J. Linn. Soc. Lond. 82: 8
-
- Arnold M. L., 1997. Natural Hybridization and Evolution, Oxford University Press, New York
-
- Barchiesi F., Cogliati M., Esposto M. C., Spreghini E., Schimizzi A. M., et al. , 2005. Comparative analysis of pathogenicity of Cryptococcus neoformans serotypes A, D and AD in murine cryptococcosis. J. Infect. 51: 10–16 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources