

Use of Cumulative Poisson Probability Distribution as an Estimator of the Recombination Rate in an Expanding Population: Example of the Macaca fascicularis Major Histocompatibility Complex
- PMID: 22384389
- PMCID: PMC3276188
- DOI: 10.1534/g3.111.001248
Use of Cumulative Poisson Probability Distribution as an Estimator of the Recombination Rate in an Expanding Population: Example of the Macaca fascicularis Major Histocompatibility Complex
Abstract
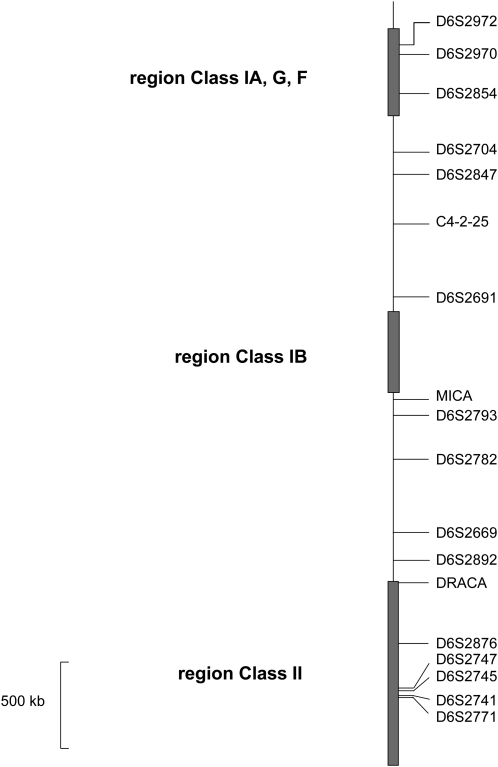
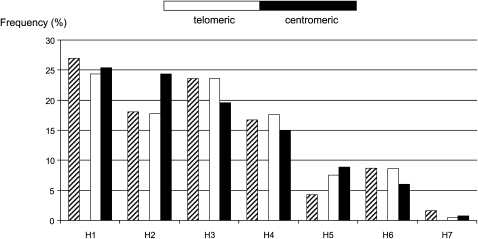
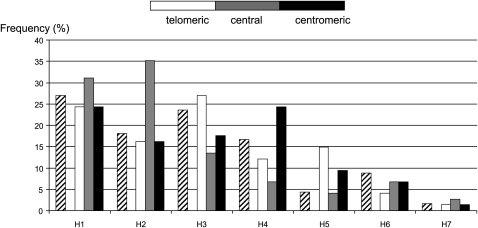
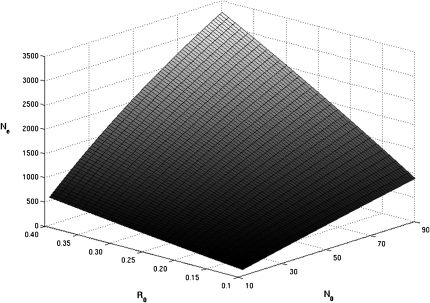
We describe a method to estimate the rate of recombination per generation from the genotypes of a large individual sample of an expanding population, for which the founding event is dated. The approach is illustrated with an application to estimating the major histocompatibility complex (MHC) recombination rate in the Mauritian macaque population. We genotyped 750 macaques by means of 17 microsatellites across the MHC region and reconstructed the seven most frequent haplotypes assumed to represent the founding haplotypes (H(rec(0))) as well as the 31% recombinant haplotypes (H(rec(h))) resulting from a variable number "h" of recombinations between the founding haplotypes. The relative frequencies of the various classes of haplotypes (H(rec(0)) and H(rec(h))) follow a Poisson distribution. By using a maximum likelihood method, we calculated the mean of the Poisson distribution that best fits the data. By dividing this mean by the number of generations (50-100) from the date of the population founding, we deduced that rate of recombination in the MHC is approximately 0.004 to 0.008 in the Mauritian macaque population. When the founding date of the population is precisely known, our method presents a useful alternative to the coalescent method.
Keywords: MHC; Macaca fascicularis; Poisson distribution; expanding population; major histocompatibility complex; recombination rate.
Figures

References
-
- Aarnink A., Dereuddre-Bosquet N., Vaslin B., Le Grand R., Winterton P., et al. , 2011. Influence of the MHC genotype on the progression of experimental SIV infection in the Mauritian cynomolgus macaque. Immunogenetics 63: 267–274 - PubMed
-
- Beck S., Trowsdale J., 2000. The human major histocompatibility complex: lessons from the DNA sequence. Annu. Rev. Genomics Hum. Genet. 1: 117–137 - PubMed
-
- Bonhomme M., Blancher A., Crouau-Roy B., 2005. Multiplexed microsatellites for rapid identification and characterization of individuals and populations of Cercopithecidae. Am. J. Primatol. 67: 385–391 - PubMed
-
- Bonhomme M., Blancher A., Cuartero S., Chikhi L., Crouau-Roy B., 2008. Origin and number of founders in an introduced insular primate: estimation from nuclear genetic data. Mol. Ecol. 17: 1009–1019 - PubMed
LinkOut - more resources
Full Text Sources
Research Materials