

Extent With Modification: Leg Patterning in the Beetle Tribolium castaneum and the Evolution of Serial Homologs
- PMID: 22384402
- PMCID: PMC3284331
- DOI: 10.1534/g3.111.001537
Extent With Modification: Leg Patterning in the Beetle Tribolium castaneum and the Evolution of Serial Homologs
Abstract
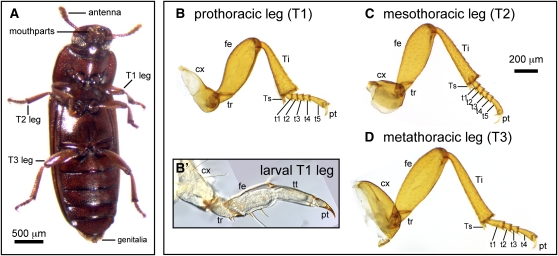
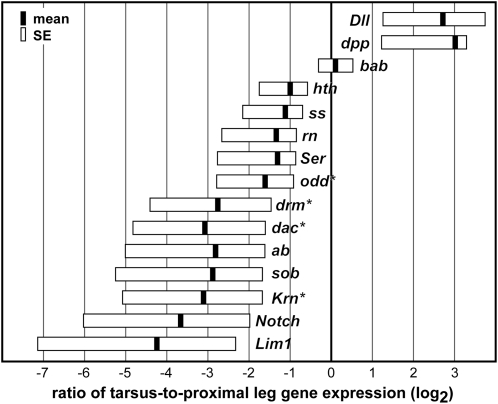
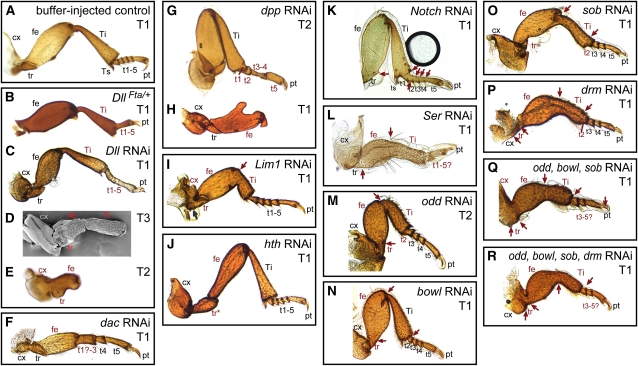
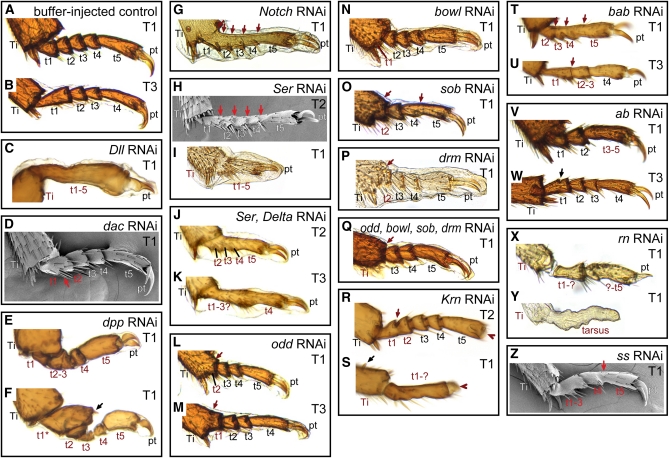
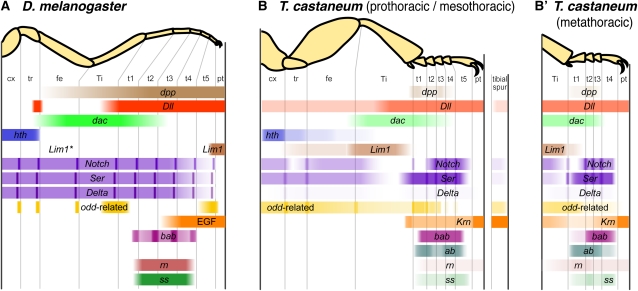
Serial homologs are similar structures that develop at different positions within a body plan. These structures share some, but not all, aspects of developmental patterning, and their evolution is thought to be constrained by shared, pleiotropic gene functions. Here we describe the functions of 17 developmental genes during metamorphic development of the legs in the red flour beetle, Tribolium castaneum. This study provides informative comparisons between appendage development in Drosophila melanogaster and T. castaneum, between embryonic and adult development in T. castaneum, and between the development of serially homologous appendages. The leg gap genes Distal-less and dachshund are conserved in function. Notch signaling, the zinc-finger transcription factors related to odd-skipped, and bric-à-brac have conserved functions in promoting joint development. homothorax knockdown alters the identity of proximal leg segments but does not reduce growth. Lim1 is required for intermediate leg development but not distal tarsus and pretarsus development as in D. melanogaster. Development of the tarsus requires decapentaplegic, rotund, spineless, abrupt, and bric-à-brac and the EGF ligand encoded by Keren. Metathoracic legs of T. castaneum have four tarsomeres, whereas other legs have five. Patterns of gene activity in the tarsus suggest that patterning in the middle of the tarsal region, not the proximal- or distal-most areas, is responsible for this difference in segment number. Through comparisons with other recent studies of T. castaneum appendage development, we test hypotheses for the modularity or interdependence of development during evolution of serial homologs.
Keywords: adult development; appendage patterning; developmental constraint; metamorphosis; serial homology.
Figures
References
-
- Abu-Shaar M., Mann R. S., 1998. Generation of multiple antagonistic domains along the proximodistal axis during Drosophila leg development. Development 125: 3821–3830 - PubMed
-
- Abzhanov A., Kaufman T. C., 2000. Homologs of Drosophila appendage genes in the patterning of arthropod limbs. Dev. Biol. 227: 673–689 - PubMed
-
- Amin A., Li Y., Finkelstein R., 1999. Hedgehog activates the EGF receptor pathway during Drosophila head development. Development 126: 2623–2630 - PubMed
-
- Angelini D. R., Kaufman T. C., 2004. Functional analyses in the hemipteran Oncopeltus fasciatus reveal conserved and derived aspects of appendage patterning in insects. Dev. Biol. 271: 306–321 - PubMed
-
- Angelini D. R., Kaufman T. C., 2005a Functional analyses in the milkweed bug Oncopeltus fasciatus (Hemiptera) support a role for Wnt signaling in body segmentation but not appendage development. Dev. Biol. 283: 409–423 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous